
Abstract
Background:
The oxidative stress hypothesis is challenging the dominant position of amyloid-β (Aβ) in the field of understanding the mechanisms of Alzheimer’s disease (AD), a complicated and untreatable neurodegenerative disease.
Objective:
The goal of the present study was to uncover the oxidative stress mechanisms causing AD, as well as the potential therapeutic targets and neuroprotective drugs against oxidative stress mechanisms.
Methods:
In this study, a systematic workflow combining pharmacological experiments and computational prediction was proposed. 222 drugs and natural products were collected first and then tested on SH-SY5Y cells to obtain phenotypic screening data on neuroprotection. The preliminary screening data were integrated with drug-target interactions (DTIs) and multi-scale biomedical data, which were analyzed with statistical tests and gene set enrichment analysis. A polypharmacology network was further constructed for investigation.
Results:
340 DTIs were matched in multiple databases, and 222 cell viability ratios were calculated for experimental compounds. We identified significant potential therapeutic targets based on oxidative stress mechanisms for AD, including NR3C1, SHBG, ESR1, PGR, and AVPR1A, which might be closely related to neuroprotective effects and pathogenesis. 50% of the top 14 enriched pathways were found to correlate with AD, such as arachidonic acid metabolism and neuroactive ligand-receptor interaction. Several approved drugs in this research were also found to exert neuroprotective effects against oxidative stress mechanisms, including beclometasone, methylprednisolone, and conivaptan.
Conclusion:
Our results indicated that NR3C1, SHBG, ESR1, PGR, and AVPR1A were promising therapeutic targets and several drugs may be repurposed from the perspective of oxidative stress and AD.
Keywords
INTRODUCTION
Oxidative stress is related to many diseases, including atherosclerosis, chronic obstructive pulmonary disease, cancer, and Alzheimer’s disease (AD), and antioxidants as small molecules have exhibited potential therapeutic effects in preclinical studies [1]. Especially, in the pathogenesis of AD, the lipids, proteins, and nucleic acids in neurons could be oxidized because of mitochondrial dysfunction, so oxidative stress plays an important role. Studies have revealed that oxidative stress is considered to cause amyloid-β (Aβ) deposition, tau hyperphosphorylation, and the subsequent death of neurons in the development of AD [2]. Mitochondrial function is significant in neurons, and recent research suggests that the restoration of mitochondrial function by antioxidant can slow the progression of AD [3]. In the past decades, many research has been conducted to investigate the impact on AD of different mitochondrial dysfunction forms, such as excessive reactive oxygen species (ROS) production, mitochondrial Ca2 + dyshomeostasis, and loss of ATP [4, 5]. It is obvious that oxidative stress plays a significant role in the formation of AD. For better understanding the mechanisms of AD and discovery of potential therapeutic targets and drugs, the oxidative stress hypothesis deserves more attention.
AD, the most common form of dementia, affects millions of people worldwide as a neurodegenerative disease, and has become one of the greatest healthcare challenges in the 21st century [6]. AD causes the death of neurons and destroys memory and thinking ability of humans, with clinical manifestations of cognitive decline and different degrees of behavioral disorders [7]. According to the 2020 International Alzheimer’s Disease Association report, the number of AD patients has reached 50 million globally and will exceed 100 million in 2050 as predicted. There are still no drugs that can slow the progression of AD during decades of research, let alone a cure [8]. However, drugs are the most important strategies to help AD patients maintain mental state, manage behavioral symptoms, and slow down the development of the disease. Currently, there are only four small molecular drugs approved by the US Food and Drug Administration (FDA) for the treatment of AD, namely, donepezil, rivastigmine, galantamine, and memantine. Memantine is an antagonist of N-methyl-D-aspartate (NMDA) receptor, whereas the other three ones are acetylcholinesterase inhibitors [9]. Yet these drugs can only be used to relieve related symptoms and cannot change the pathogenesis of AD. Aduhelm was approved by the FDA in 2021 as a biomacromolecular drug; it is found to reduce Aβ deposition in the brains of AD patients. However, its efficacy in a wider range of patients remains to be confirmed [10]. 121 anti-AD drug candidates have been investigated in clinical trials in the past five years, but the drug failure ratio reached to 98.7% [11]. The main reason for such a situation is that the multiplex pathogenesis of AD remains unclear. Researchers have discovered in the past decades that the pathological features are extracellular Aβ plaques and neurofibrillary tangles in neurons caused by hyperphosphorylation of tau protein, accompanied by the loss of central cholinergic neurons and proliferation of glial cells [12]. Many hypothetical pathogenic mechanisms have been proposed, such as the Aβ hypothesis [13], tau hypothesis [14], neuroinflammation [15], and free radicals and oxidative stress [16]. Most of the drugs are developed based on these hypotheses and ended in failure; the FDA has not approved any new small molecular anti-AD drugs since 2003. Although Aβ is still the most investigated target, non-amyloid hypothesis such as neuroinflammation and oxidative stress related targets are getting more researchable. Evidence has shown that AD is a progressive and complex disease involving multiple biochemical processes beyond the accumulation of Aβ and tau [17]. Besides, more studies have focused on the oxidative stress and mitochondrial dysfunction in AD and the research results are promising [18].
Novel drugs development and mechanisms analysis for AD are becoming more difficult to move forward than ever. Computational systems pharmacology methods, based on network pharmacology and multiple computational tools to realize multi-stage, multi-level, multi-space analysis on the interactions between drugs and disease through an integrated system, may help accelerate the process. Experimental-computational approaches have been developed and applied to find new therapeutic targets and to repurpose approved drugs [19]. Chemogenomics-knowledgebase systems pharmacology methods had been applied to discover cannabidiol-related targets and potential links to neurological diseases [20]. Wang et al. used computational systems pharmacology methods to discover the effective chemical components in herbal formula Tian-Ma-Gou-Teng-Yin against AD and found some targets potentially related to AD [21]. Another systems pharmacology study based on collected data and computational analysis found the antiplatelet and neuroprotective mechanisms in the treatment of ischemic stroke of a traditional Chinese medicine Deng-Zhan-Xi-Xin injection, which provides a systems perspective to study the relevance between herbal drugs and disease processes [22]. Based on an endophenotype-based methodology and large-scale data, researchers identified sildenafil as a potential disease risk modifier for AD [23]. But the relationship between sildenafil and AD still needs more randomized controlled trials. These studies have limitations in data acquisition and analysis to some extent, this may lead to the problems of obtaining useful data for and restrictive results. Therefore, multi-omics data, real-world experimental data and systematic workflow are essential for uncovering potential pathogenic mechanisms and discovering effective drugs for AD.
In view of the significance of oxidative stress mechanisms and the complexity of AD, we should move beyond the “one drug ⟶ one target ⟶ one disease” paradigm and explore the “multiple drugs ⟶ multiple targets ⟶ multiple diseases” possibility to discover novel strategy [24]. In this study, we proposed a computational systems pharmacology workflow based on oxidative stress hypothesis, represented in Fig. 1, to uncover potential therapeutic targets and pathogenic mechanisms of AD. We first conducted pharmacological experiments on 222 drugs and drug-like compounds on nerve cells to obtain preliminary phenotypic screening data. And then integrated DTIs information and multi-scale biomedical data, combining methods of statistical tests, biological pathways enrichment analysis, and construction of polypharmacology networks for further analysis. Finally, we identified a couple of potential therapeutic targets for AD, such as NR3C1, SHBG, ESR1, PGR, and AVPR1A. These target genes can closely interact with neuroprotective effects and the pathogenesis of AD, especially the oxidative stress mechanisms. Some FDA approved drugs used in this research were also found to exert neuroprotective effects against AD, such as beclometasone, methylprednisolone, and conivaptan. In the enrichment analysis of drugs with significant neuroprotective effects, 50% of the top 14 pathways were found to closely associated with AD, such as arachidonic acid metabolism, serotonergic synapse, and neuroactive ligand-receptor interaction. In general, we employed a systematic method for investigating potential mechanisms and therapeutic targets for AD. This workflow could also provide a holistic perspective to discover potential pathogenesis for other diseases with complex pathogenic mechanisms.
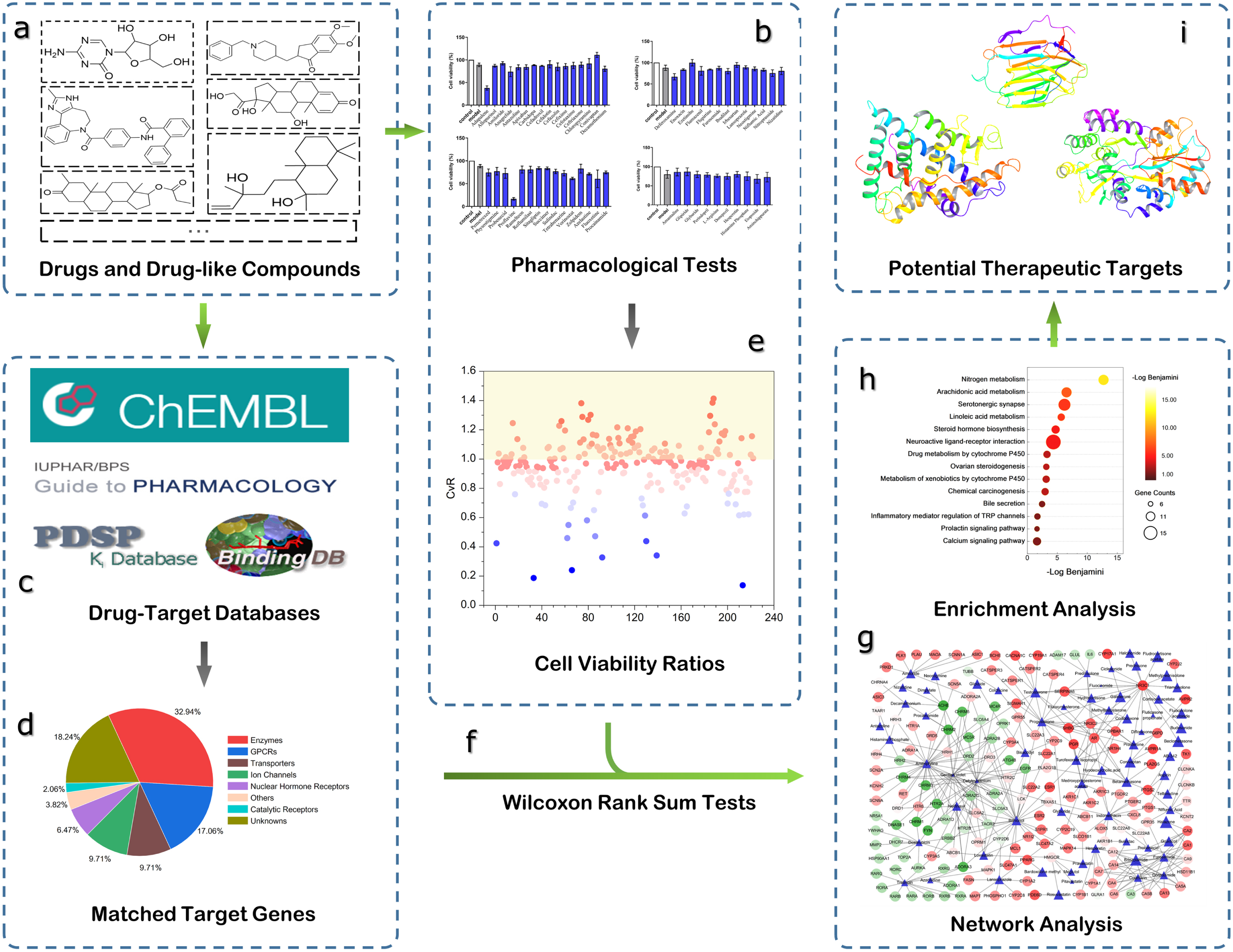
The workflow of the computational systems pharmacology approaches used in this study, including (a) selection of drugs and drug-like compounds for this study, (b) pharmacological experiments for 222 drugs on SH-SY5Y cells, (c) matching targets for all drugs from multiple databases, (d) types of matched targets according to IUPHAR Guide to PHARMACOLOGY classification, (e) calculation of experimental cell viability ratios for every tested drug, (f) Wilcoxon rank sum tests for matched target genes, (g) construction of DTI networks for further analysis, (h) GSEA analysis for drugs showing neuroprotective effects, (i) selection of potential therapeutic targets for AD.
MATERIALS AND METHODS
Data collection and preparation
Due to the complexities and internal relationships of the hypothetical mechanisms of AD, a number of drugs which had potential correlation with several AD hypotheses were collected from DrugBank [25], including not only drugs related to neuroinflammation and other inflammations, acetylcholinesterase, blood vessels and pressure, but also some antibacterial drugs related to gut flora. Since the relationship between gut and AD has been reported more frequently [26]. Additionally, some natural products from TargetMol and SPECS databases were collected to help extend the chemical space of our data set and provide a broader scope for further pharmacological experiments and computational analysis. Other information about these drugs and natural products were also collected, such as SMILES, molecular weight (MW) and Log P. All these chemical structures were converted into canonical SMILES format by Open Babel toolkit (version 2.3.1). All the compounds were purchased from TargetMol and SPECS and divided into several groups for biological tests.
Neuroprotective activity against oxidative damage induced by H2O2 on SH-SY5Y cells
Pharmacological experiments were conducted for collected compounds to explore their protective effects on H2O2 induced neurotoxicity. Cell viability was determined by MTT assay. SH-SY5Y cells were seeded into 96-well plates. After 24 h, tested compounds were added at 10μM with experimental groups. After 2 h, all cells were exposed to H2O2 (400μM) for 30 min. After another 4 h of incubation at 37°C, supernatants were removed and formazan particles were dissolved in DMSO. The absorbance was measured using a Synergy 2 Multi-Mode Microplate Reader (BioTek, Winooski, VT) at the wavelength of 490 nm. Calculation formula is as following equation:
Cell viability ratio (CvR) was proposed to enable the direct comparison and analysis of cell viability for different compounds in different test groups. The definition of CvR is as follows:
Targets matching for experimental compounds
Experimentally determined DTIs were collected from four public databases, including ChEMBL [27] (version 29), BindingDB [28] (accessed in April, 2021), IUPHAR Guide to PHARMACOLOGY [29] (accessed in April, 2021), and PDSP Ki Database [30] (accessed in April, 2021). Firstly, the Open Babel toolkit (version 2.3.1) was used to convert chemical structures of all compounds into canonical SMILES format. For each of the compounds, the corresponding targets were then matched from these databases under the following three criteria by python scripts: (i) target proteins are from Homo sapiens and have unique UniProt [31] accession numbers; (ii) target proteins were labelled as “reviewed” in the UniProt knowledgebase; (iii) IC50, EC50, Ki, or Kd values ≤10μM. For one target, if the above three criteria were met, this DTI data will be collected. Duplicated DTIs were then removed. Finally, official gene symbols of targets were obtained from the HUGO Gene Nomenclature Committee (HGNC) [32], according to their UniProt accession numbers. The prepared DTIs and related information were used in the subsequent computational prediction and analysis.
Statistical analysis
Wilcoxon rank sum test, also known as Wilcoxon-Mann-Whitney test, was implemented as the nonparametric statistical method. It was to determine whether two distribution columns were significantly different [33] by comparison of two non-parametric group data or two independent samples. Finally compared the null hypothesis to alternative hypothesis for differences or similarities.
In this study, two sets Xj and Yj were defined for each matched target Tj of the selected drugs. Xj is the set containing the CvR values of the drugs that have known interactions with Tj, while Yj is the set containing the CvR values of the drugs that do not have known interactions with Tj. Mathematically, assuming that D is the set containing all the collected drugs, Lj is the set of known ligands of target Tj, and f (D
i
) is the CvR value of drug Di, these two sets can be represented as:
Network visualization
DTI networks were constructed based on the following information: (i) the known existing DTIs; (ii) the p value of each statistically tested gene; (iii) the CvR of each pharmacologically tested drug. Cytoscape [34] (version 3.8.1) was used to construct DTI networks with weighted nodes for visually analyzing the comprehensive quantitative interaction between drugs and target genes. Nodes of drugs and targets in DTI networks were labeled by different colors and shapes according to the attribute data for further network analysis. NetworkAnalyzer, a built-in module of Cytoscape, was then used to calculate the network distribution topological information, including nodes degree, neighborhood connectivity, average shortest path length and closeness centrality. Morgan fingerprints and chemical similarities of all compounds were calculated by RDKit, an open-source cheminformatics package.
Biological function enrichment analysis of predicted genes
Gene set enrichment analysis was conducted for these matched drug targets to detect key targets and pathways closely related to neuroprotection by using the Database for Annotation, Visualization and Integrated Discovery (DAVID, version 6.8), a web server that provides a comprehensive set of functional annotation tools for investigators to understand biological meaning behind large list of genes [35]. Pathway enrichment analysis were performed on the corresponding target gene sets of drugs with the pharmacological test results of CvR >1. Significant pathways with potential neuroprotective effects for oxidative stress and AD were found by biologically analyzing the results of KEGG pathway enrichment analysis.
RESULTS
Data collection and phenotypic screening experiments
Information of several drugs and drug-like compounds used in pharmacological experiments with different indications
A total of 222 drugs and natural products were collected from DrugBank, TargetMol, and SPECS databases. Among them, there were 77 natural products and 145 small molecular drugs. These compounds were divided into different groups to perform pharmacological tests on H2O2 induced SH-SY5Y cells for obtaining the preliminary screening data. Some compounds were the FDA approved drugs. Table 1 showed some selected compounds and corresponding information, including names, indications, chemical formulas, Chemical Abstract Services Registry Numbers (CASRN), structures and some specific external links. All drugs have a very wide range of indications. See more detailed information about 222 drugs and drug-like compounds in Supplementary Table 1.
222 compounds were labeled and divided into several groups to make the experiments more efficient. The pharmacological test results of cell viability of these compounds were obtained as the fundamental dataset for the following computational studies. All cell viability results of the tested compounds were also put in Supplementary Table 1. Herein, the pharmacological test results of 54 compounds were shown in Fig. 2A to present the neuroprotective abilities and neurotoxic effects of tested compounds. Cell activities of compounds higher than corresponding model groups indicated potential neuroprotective effects. For example, Conivaptan, with a cell viability of 110.977%, showed neuroprotective effects on H2O2 induced SH-SY5Y cells. Cell activities of compounds lower than corresponding model groups indicated potential toxicity to nerve cells. Adapalene did not show neuroprotective effects, on the contrary, it showed neurotoxicity with a cell viability of 37.798%. In order to present the quantitative measurements of neuroprotective abilities of compounds, CvRs were calculated to compare and analyze cell viabilities of these compounds in different test groups directly. The scatter information and distribution of CvRs of all compounds were shown in Fig. 2B, most of the CvRs were scattered between 0.1 and 1.4, indicating that the tested compounds had different biological effects and wide threshold range. Compounds with CvRs greater than 1.0 indicated potential neuroprotective effects on H2O2 induced SH-SY5Y cells, while CvRs lower than 1.0 indicated compounds did not show neuroprotective effects, some of them even showed the tendency of neurotoxicity. Figure 2C showed the frequency of CvR was distributed around 1.0, 77.5% of all CvRs were between 0.8 and 1.2. 88 compounds with CvRs >1.0 and the maximum CvR reached 1.413, 134 drugs with CvRs <1.0 and the minimum CvR reached 0.137. Considering the comprehensive CvR distribution, these compounds with higher CvRs exhibited more research significance.
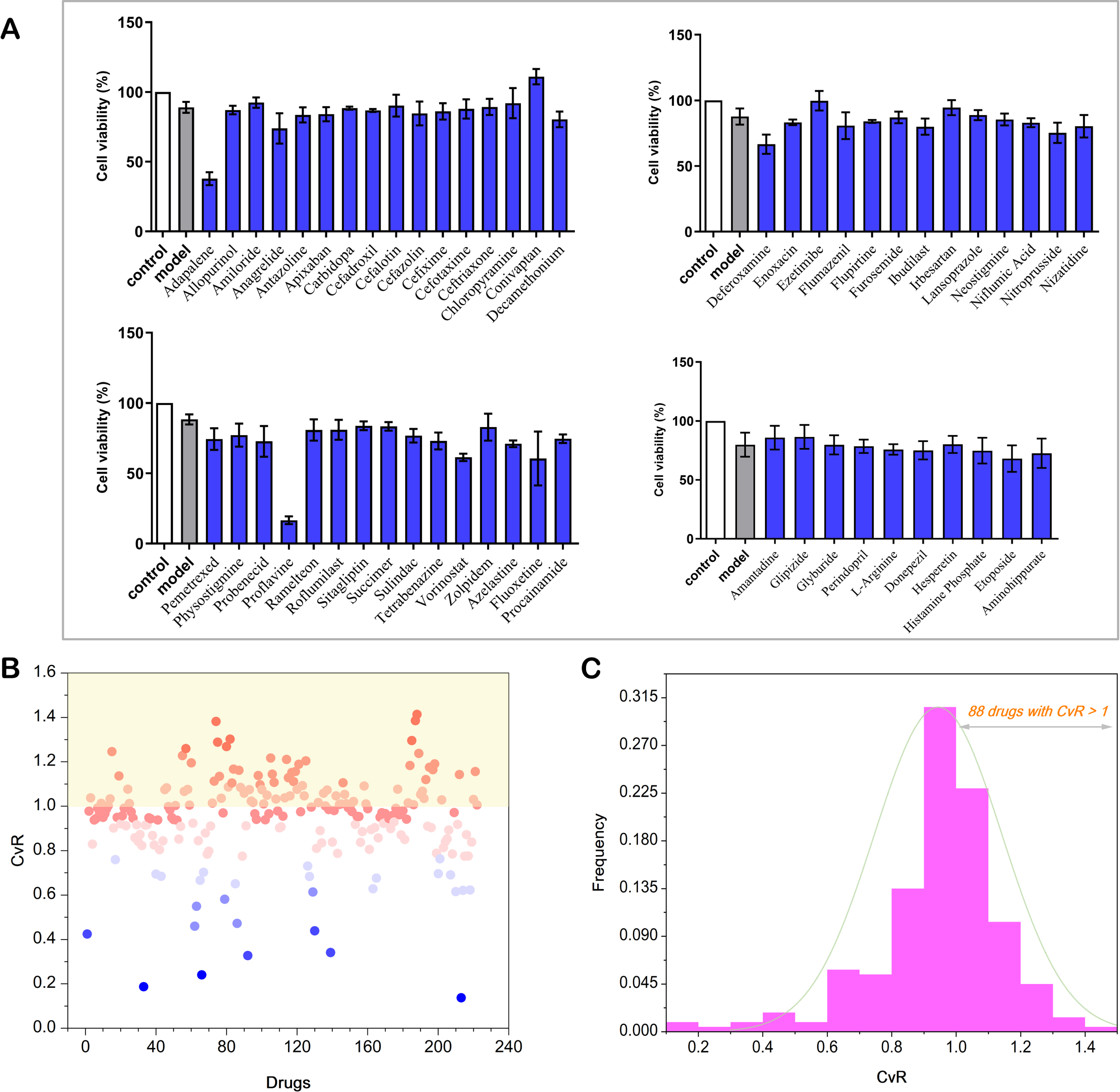
Cell viability results of CvRs. A) The experimental cell viability results of 58 compounds tested on H2O2 induced SH-SY5Y cells. B) The overall scatter diagram of all drugs based on their CvRs. C) The frequency histogram distribution of all drugs (88 drugs with CvRs >1).
Target matching for experimental compounds
All the canonical SMILES formats of the tested compounds were obtained by Open Babel toolkit, then existing known target genes for them were matched from four databases (ChEMBL, BindingDB, IUPHAR Guide to PHARMACOLOGY, and PDSP Ki Database). After obtaining experimentally verified target genes, 130 compounds matched to at least one target gene and a total of 340 genes were matched out for these compounds. The average number of matched genes for each compound was approximately 2.62. As shown in Fig. 3A, G protein-coupled receptors (GPCRs) and enzymes accounted for half of the matched targets, transporters and ion channels both accounted for approximately 9.71% and catalytic receptors accounted for 2%. These matched genes included those had known active interactions with these compounds and those had known inactive interactions. Therefore, the following statistical analysis could be better performed based on the diversity of the number of genes. Matching results showed that 91 compounds had at least 2 genes and 29 compounds had at least 20 genes. Especially, 35 known genes were matched out for Amitriptyline, 34 genes for Indomethacin and 26 genes for Vorinostat. Detailed target matching information was put in Supplementary Table 2.
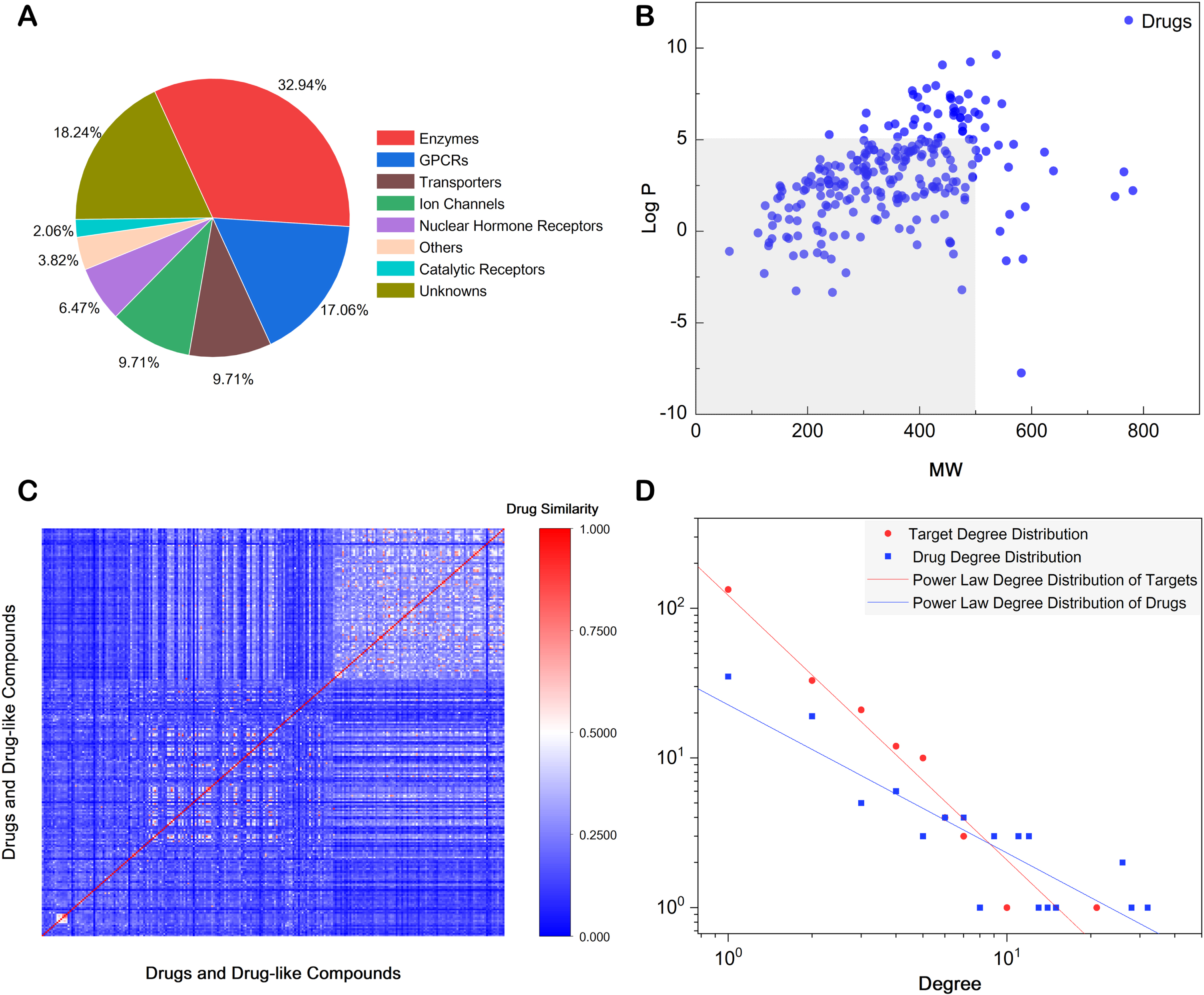
Property analysis of drugs and DTI networks. A) Percentage of different kinds of matched targets according to IUPHAR classification. B) Chemical space distribution of drugs using in pharmacological experiments, depicted by two descriptors: MW and LogP. C) Chemical similarities of all drugs based on Morgan fingerprints. D) Power-law degree distribution of two kinds of nodes in DTI networks: drugs (blue), targets (red).
Statistical tests for matched genes
Wilcoxon rank sum test was performed for all the 340 genes based on the CvRs of all compounds obtained by pharmacological experiments. Wilcoxon test results for 220 genes were obtained by R, while the statistical results of 120 genes could not be obtained, because these genes had no known interactions with the compounds. For one gene, taking ABCB1 as an example, there were three compounds with experimentally determined interactions with ABCB1 and CvRs of these compounds were 1.0075, 0.7879, and 0.9809 respectively. Therefore, the three CvRs were used as Xj for statistical test. The CvRs of the rest compounds were used as Yj to ensure the total number of compounds used in one statistical test were consistent. If the p value of the statistical test was greater than the set critical value, null hypothesis H0 was accepted and alternative hypothesis H1 was rejected, indicating there was no significant relationship between ABCB1 and neuroprotection; otherwise, null hypothesis H0 was rejected and alternative hypothesis H1 was accepted, indicating ABCB1 was significantly related to neuroprotection. Actually, the statistical test result p value of ABCB1 was 0.7023, presenting that ABCB1 had no statistically significant relationship with neuroprotection. All statistical tests for aforementioned genes were based on the above hypotheses to conduct, statistical test results of 220 genes were then obtained. Then the genes were ranked by corresponding p value. The overall distribution of p values of the matched genes was shown in Fig. 4A; the horizontal axis represents each gene and the vertical axis represents the corresponding p values. Genes were ranked and colored by p values, and colors close to dark representing lower p values. Among these genes, NR3C1 was ranked first with p value of 0.0287. The detailed statistical test results were put in Supplementary Table 3.
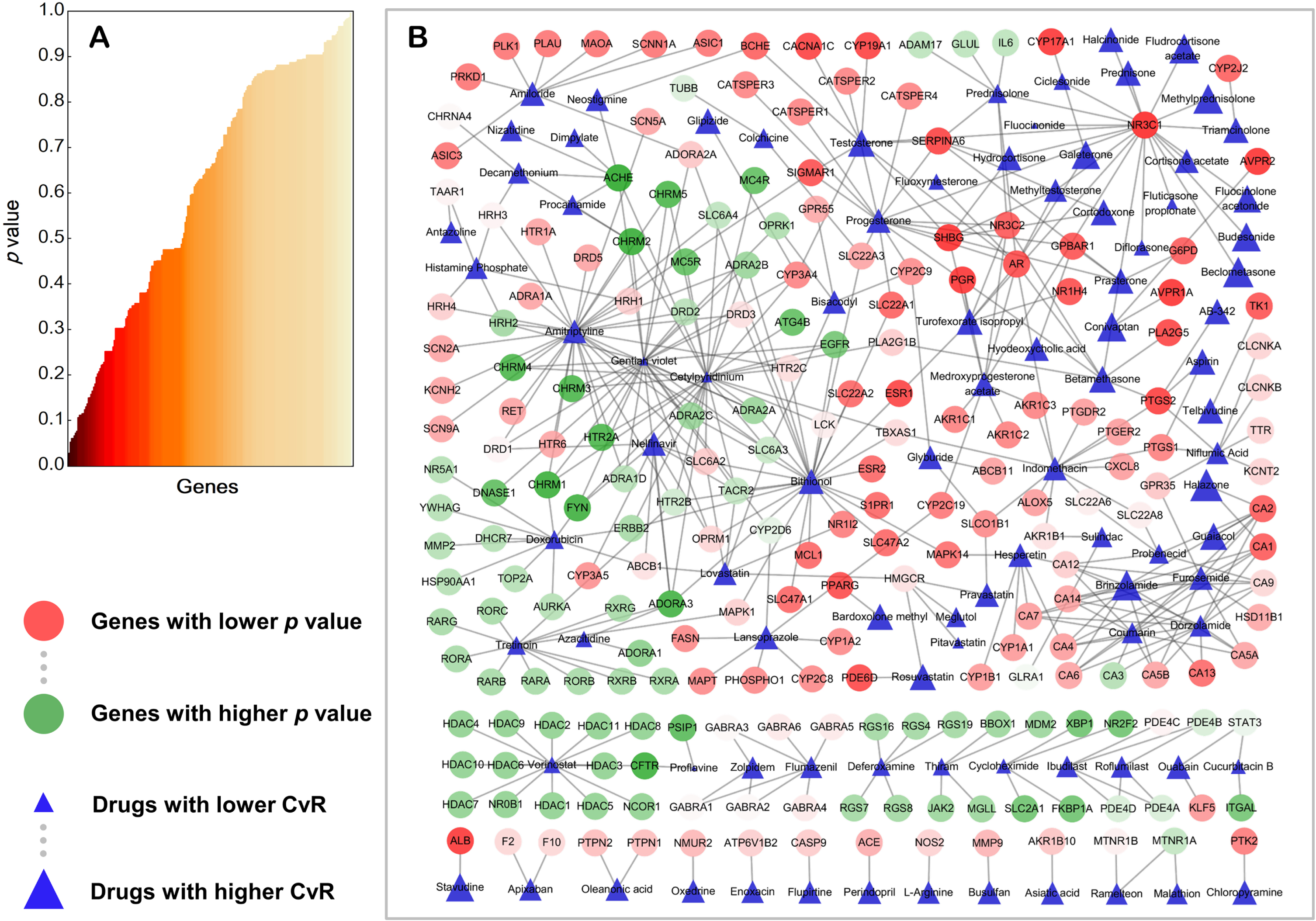
The p values distribution of genes and DTI networks with weighted nodes. A) Distribution of p values for target genes of experimental drugs. B) DTI networks. Target genes are represented by circles with different degrees of colors, red: lower p values, green: higher p values. Drugs are represented by blue triangles with different sizes determined by CvRs, larger: higher CvRs, smaller: lower CvRs. This figure was plotted using Cytoscape v3.8.1.
Drug-target network construction and visualization
The relationships between target genes and corresponding drugs, and the relationships between target genes and potential neuroprotective effects were then analyzed through the bipartite networks, which were constructed based on CvR and statistical test p values. Cytoscape was used to construct the weighted bipartite networks for visual analysis. In the constructed network, there were two types of nodes, drug nodes and target gene nodes, and edges between the two different sets of nodes. One target gene node was connected to one drug node by an edge under the condition that the gene was the actual target of the drug. There were no edges under the condition that no clear pharmacological interactions were shown between the two types of nodes. In this way, a bipartite network was obtained via a set of target genes and drugs. As shown in Fig. 3B, the chemical space distribution of drugs using in pharmacological experiments were depicted by two descriptors, MW and Log P. Distribution of MW of these compounds was between 0 and 800, distribution of Log P was from -10 to 10, more than half of the compounds met Lipinski’s rule of five [36]. Chemical similarities of all drugs based on Morgan fingerprints were also calculated by RDKit package. Figure 3C showed an average Tanimoto coefficient of 0.19, indicating the structural diversity among the compounds in the network. The nodes of target genes were colored based on the p values, the shades and dimensions of drug nodes were determined according to the CvR of drugs. DTI networks were constructed to highlight significant genes and drugs closely related to neuroprotective effects and AD. Degree distribution of two kinds of nodes in the network were described by the power-law degree distribution. The results were calculated by NetworkAnalyzer and shown in Fig. 3D. Red circles and blue squares represented target degree distribution and drug degree distribution respectively. The fitted red line and blue line described the power-law degree distribution of targets and drugs respectively, demonstrated that the networks of targets and drugs are scale-free networks. Other topological information of DTI networks was put in Supplementary Table 4.
These circular nodes in DTI networks, as shown in Fig. 4B, represented target genes. Colors for target nodes were represented by gradient color from green to red. The closer the color of circular nodes approached to green, the greater were the p values of the genes. This reflected these genes had relatively lower correlations with neuroprotection. Yet, color of target nodes approaching to red represented the smaller p values of the genes, indicated that these genes had relatively higher correlations with neuroprotection and AD. This type of genes could have that potential to become therapeutic targets in the process of neuronal cell death in neurodegenerative diseases. The drug nodes were represented by blue rectangles. Smaller area and lighter color of the rectangles represented lower CvR of the drugs, indicating that these drugs could have no significant relationships with neuroprotection. Conversely, larger area and darker color of the rectangles represented higher CvR, indicated these drugs had more significant relationships with neuroprotective effects against neurotoxicity. From Fig. 4B, it is easy to see those genes such as NR3C1, SHBG, PTGS2, ESR1, and BCHE were found at key positions in DTI networks and deeper in red. These genes connected to several dark blue nodes of drugs simultaneously, indicating that these genes were inseparable from neuroprotective effects in nerve cells. As shown in DTI networks, AVPR1A also presented correlations with potential neuroprotection as a gene encoding arginine vasopressin receptor 1A, which belongs to a subfamily of G protein-coupled receptors. Therefore, in our study, NR3C1, SHBG, ESR1, PGR, and AVPR1A were suggested as potential targets for AD. In addition, some experimental evidence to support our results will be further discussed.
Gene set enrichment analysis
Enrichment analysis of biological processes, pathways, or molecular functions of selected AD related gene sets could help understand the key functions of genes and increase insights into the underlying mechanisms of the gene sets of interest [37]. The results of the enrichment analysis are shown in Supplementary Table 5. Benjamini is the correction value for the p value in the enrichment results, represented by PBen in this study. The smaller the PBen is, the higher is the credibility of the enriched pathway, which means the top ranked pathways are more relevant to neuroprotection and AD. The enrichment pathways were ranked according to -log PBen obtained by the enrichment analysis. The top 15 pathways related to AD were shown in Fig. 5, including nitrogen metabolism (PBen = 1.93×10–13), arachidonic acid metabolism (PBen = 2.68×10–7), serotonergic synapse (PBen = 6.39×10–7), linoleic acid metabolism (PBen = 2.20×10–6), steroid hormone biosynthesis (PBen = 1.88×10–5), neuroactive ligand-receptor interaction (PBen = 4.55×10–5), inflammatory mediator regulation of TRP channels (PBen = 0.0205), calcium signaling pathway (PBen = 0.0241), and other neuroprotection related pathways. The size of the circle area indicated the number of genes, 16 genes enriched in the neuroactive ligand-receptor interaction pathway were the most, 6 genes enriched in the Prolactin signaling pathway were the least.

Top 14 enriched pathways in enrichment analysis results using DAVID v6.8. The circles are colored according to -logPBen, the color approaching to yellow representing lower values of PBen and higher values of -logPBen indicating the higher significance of pathways. The sizes of circles are determined by the counts of genes included in each pathway.
Among top 15 pathways, arachidonic acid metabolism is closely related to AD. Studies had revealed that arachidonic acid and other unsaturated fatty acids were significantly dysregulated in the brains of patients with different processes of AD [38]. It was found in previous studies that serotonergic synapse was correlated with neurotransmitter transmission in brain neurons of AD patients, the reduction of 5-HT reuptake transporters (SERTs) caused the imbalance of neurotransmitter transmission in AD, which is possible to aggravate related phenotypic symptoms in AD patients [39]. There were correlations between neuroactive ligand-receptor interaction pathway and AD according to the previous analysis of the gene expression network in the blood of AD patients and healthy people. AD modules related to the neuroactive ligand-receptor interaction pathway in the network did not appear in gene expression network of healthy people, suggested genes in this pathway might be significant targets for AD mechanism research and drug discovery [40].
A number of studies had shown that Transient Receptor Potential (TRP) channels participated in the process of AD through the mechanism of Ca2 + regulation, oxidative stress and the production of inflammatory mediators. In particular, neuroinflammation has always been as the key mechanism involved in neurodegeneration. TRPV1, a member of TRP family, acts as an essential role in neuroinflammation. Recent studies demonstrated that TRPV1 could be a therapeutic target to alleviate the progress of AD [41]. In AD models, researchers found that TRPC6 regulates the cleavage of amyloid-β protein precursor (AβPP) protein through γ-secretase. Therefore, TRPC6 might be a new strategy to reduce the formation of Aβ in the process of AD [42]. This evidence showed that the inflammatory mediator regulation of TRP channels pathway would be significantly useful to deal with the phenotypic symptoms in AD. Multiple targets in this pathway have the potential to become effective targets.
Calcium signaling pathway is also closely related to AD. Ca2 + is a key part for normal physiological functions of neurons such as neuronal survival, growth, and differentiation. There was strong evidence suggesting that abnormal Ca2 + levels could result in the imbalance of intracellular signals and consequently cause neuronal death [43]. The destruction of Ca2 + homeostasis in neurons leads to the generation and accumulation of reactive oxygen species (ROS), then caused mitochondrial dysfunction in neurodegenerative diseases. Studies showed that the increase of Ca2 + closely related to other pathogenic mechanisms of AD, such as Aβ deposition [44], the imbalance between ROS and antioxidant function [45] and mitochondrial dysfunction in neurons [46]. The relationships among oxidative stress, mitochondrial dysfunction, and the pathogenesis of AD were connected with the increasing intracellular Ca2 + levels. The dysregulation of Ca2 + signals was one of the key processes of early neuron loss in AD. Researchers have not yet found the original sources and mediators of Ca2 + increase. However, it is still promising to prevent excessive neuronal death in the early stages of AD by developing pathway-specific drugs targeting the genes involved in the calcium signaling pathway.
DISCUSSION
In this study, we developed a computational systems pharmacology workflow combining pharmacological oxidative experiment screening data, for identification of potential oxidative stress mechanisms and therapeutic targets for AD. As the research results showed, we uncovered some potential therapeutic targets for AD from the perspective of oxidative stress hypothesis and suggested several old drugs might be repurposed.
Discovery of potential therapeutic targets
Several target proteins, including NR3C1, SHBG, ESR1, PGR, and AVPR1A were suggested as the potential targets for AD based on our experimental results and computational analysis.
From Fig. 4B, we could see that the subnetwork around NR3C1 was covered by s set of deep blue nodes with significant neuroprotective effects, indicating multiple drugs exert neuroprotective effects through NR3C1 gene. NR3C1, the encoding gene of glucocorticoid receptor (GR), is the target of glucocorticoids. It is a transcription factor as well as a regulator of other transcription factors, after binding with a specific ligand, NR3C1 will be transported into the nucleus to participate in some biological processes such as cellular inflammation and apoptosis [47]. Human and animal model biological assays showed that high levels of glucocorticoids could act on GR to cause cognitive impairment, Aβ aggregation, tau hyperphosphorylation, neuroexcitatory toxicity and neuroinflammation and other processes, finally promoted the occurrence and development of AD [48]. Therefore, some therapeutic strategies that directly target GR to reduce its activity had been proposed and tested. These methods became particularly helpful in the early treatment of AD patients [49]. Small molecular compounds targeting AD by reducing the activity of GR had been proposed in the past decades, including corticosterone, AL-438 [50], CORT108297, and CORT113176 [51]. These compounds could reduce AD-related symptoms. However, they may have unexpected side effects on human bodies due to the influences of the ubiquitous expression of GR.
Several drugs were closely related to anti-inflammatory effects in the DTI networks, such as beclomethasone, fludrocortisone acetate, and budesonide. These drugs were possible to exert anti-inflammatory effects by targeting NR3C1 to reduce neurotoxicity, eventually produced neuroprotective effects. NR3C1 has the potential to become a promising therapeutic target for neuroprotection against oxidative stress hypothesis and AD. NR3C2 and NR1H4 also showed significant potential in neuroprotection, implying nuclear receptor superfamily might play an important role.
ESR1 is a gene encoding the estrogen receptor α (ERα), exhibiting potentially important neuroprotective effects in DTI networks as shown in Fig. 4B. Estrogen has already presented anti-inflammatory, anti-oxidant and effects, and showed certain neuroprotective effects in previous studies [52]. This corresponds to the results of our oxidative stress experiments. The neuroprotective effect of estrogen in AD is that estrogen can affect the hydrolysis process of AβPP. Studies revealed that estrogen could up-regulate α-secretase and γ-secretase and downregulate the expression of β-secretase simultaneously, then promoted AβPP production during the culture of human fetal neurons and neuroglia cells [53]. In aromatase gene knockout AD mouse models, due to the inability to synthesize estrogen, β-secretase increased and acted on the β site of AβPP to form Aβ42, accelerating the deposition of Aβ and the formation of amyloid plaques and leading to early AD symptoms in mice [54]. By acting on pS214, tau-1, pS396, and pS404, estrogen could also inhibit the activation of protein kinase A (PKA) to reduce the hyperphosphorylation of tau and eventually slow down the development of the pathological process of AD [55]. In addition, studies showed that estrogen could exert neuroprotective effects via antagonizing the neurotoxicity induced by glutamate and inhibiting cell apoptosis [56, 57]. All evidence indicated that estrogen and ERs play vital roles in the occurrence and development of AD. Combining our experimental and computational results, ESR1 closely relate to the pathogenesis and neuroprotection not only through the oxidative stress hypothesis, but other hypotheses of AD. ER modulators might be effective druggable targets to accelerate anti-AD drugs development.
As shown in Fig. 4B, PGR also shows potential neuroprotection effects in the DTI networks and bioinformatics analysis. The experimental study of Wu et al. found that progesterone could increase the expression level of GLUT3 and GLUT4 in AD mice models induced by Aβ and improve learning and memory ability of mice [58]. SHBG also exhibits potential neuroprotective effects in the DTI networks as a gene encoding sex hormone binding globulin. The previous studies conducted by Xu et al. on AD patients and control groups showed that plasma SHBG levels in AD patients were higher than those in control groups. Preliminary experimental evidence of the upregulation of plasma SHBG levels indicated that bioavailability of SHBG and functional hormones in plasma may be related to the pathogenesis of AD [59]. Therefore, higher SHBG levels may be risk factors for cognitive dysfunction, which might be one of the reasons for memory loss in elderly patients with AD. The direct mechanical relationships between SHBG and cognitive dysfunction in AD patients, however, have not been illustrated [60]. Herein, SHBG and sex hormone receptors are significant target genes and have that potential to be therapeutically effective targets for novel drugs development as part of the potential pathogenesis of AD.
AVPR1A, together with AVPR2, present correlations with potential neuroprotection effects in DTI networks. Experimental studies have shown that, in Aβ25–35-induced AD animal models, intraventricular injection of AVP at an appropriate concentration could enhance the spatial learning ability of AD mice and effectively prevent behavioral disorders caused Aβ25–35 [61]. Studies also revealed the positive effects of AVP in APP/PS1-AD mice models, demonstrating that the intranasal administration of AVP could effectively improve the working memory and long-term spatial memory of APP/PS1-AD mice [62]. These studies demonstrated that the changes in central nervous system AVP were inextricably related to the occurrence and development of AD. Changes of the concentration of AVP might have an impact on some AD related phenotypic symptoms. Therefore, there is a strong implication that AVPR1A and AVPR2 could be potential therapeutic targets in the pathogenesis of oxidative stress and AD.
As expected, BCHE shows a strong correlation with neuroprotection in the DTI networks. As a butyrylcholinesterase, evidence showed the close relationships with AD-related pathways, especially cholinergic hypothesis. Studies illustrated that BCHE had a greater impact on AD related phenotypes and BCHE modulators might have a potential impact on the remission of neurodegenerative diseases [63]. Three of the 4 small molecule drugs approved by the FDA are cholinesterase inhibitors, so BCHE also has the potential to be one of the relevant therapeutic targets against AD.
In addition to the genes mentioned above, other genes, such as TTR, CACNA1C, PTGS2, PLA2G5, and PED6D in the DTI networks, are also potentially related to neuroprotection effects and the occurrence and development of AD. These target proteins might also worth investigating in the process of targets identification and anti-AD drugs development.
Discovery of drugs with potential therapeutic effects against oxidative stress and AD
Among the old drugs tested in pharmacological oxidative experiments, beclometasone (CvR = 1.302), methylprednisolone (CvR = 1.268), fludrocortisone acetate (CvR = 1.259), and budesonide (CvR = 1.228) targeted the NR3C1 gene with high CvR, indicating that these drugs could present good neuroprotective effects. Moreover, anti-inflammatory effects could be produced by these drugs.
Inflammation, one of the significant pathogenic hypotheses of AD, can cause a variety of central nervous system diseases including AD directly or indirectly. Previous studies revealed that a number of proinflammatory genes would be expressed in the brains of AD patients. Impairments of memory and neurodegeneration in the hippocampus of AD rat models were mediated by Cyclooxygenase-2 induced neuroinflammation [64]. Experiments showed that the synthetic glucocorticoid budesonide and Cyclooxygenase-2 activity related inhibitors had synergistic effects in regulating the level of inflammation, indicated that budesonide might provide potential therapeutic effects in the design of treatment drugs for complex neuroinflammatory symptoms involved in AD [65]. A study also showed by inhaling budesonide in mice with neuroinflammation, the levels of inflammatory factors in the brains of mice could be downregulated, and the neuroinflammation in the brains of mice could be reduced [66]. Studies also found among the available anti-inflammatory drugs, glucocorticoids could produce significant neuroprotective in the treatment of AD. In encephalomyelitis mice model experiments, methylprednisolone encapsulated in glutathione pegylated liposomes exerted significant therapeutic effects on central inflammation, suggesting another potential treatment strategy for brain neuroinflammation in AD patients [67]. In neuroinflammation and neurodegenerative diseases mice models, methylprednisolone encapsulated in GSH-PEG liposomes or a lower concentration of methylprednisolone could significantly reduce the amount of astrocyte activation and the loss of axons of the neurons in central nervous system [68].
Testosterone (CvR = 1.1442) also presents potential neuroprotective and anti-AD effects in our study. Testosterone has many targets in the DTI networks, including BCHE, AR, NR3C1, CACNA1C, and SIGMAR1. Among them, BCHE and NR3C1 are closely related to neuroprotection and AD in our analysis results. Lower p values of these genes reflect their significant neuroprotective effects. So, testosterone could exert its potential effects in coping with AD related symptoms. Studies revealed that testosterone levels are related to protective measure and can be used as one of the diagnostic factors to prevent the occurrence of male AD in the future [69]. The decrease levels of testosterone in the serum are related to normal male aging, causing a series of diseases, and the lower testosterone levels in the brain increase the risk of AD. Although it is unclear whether testosterone depletion leads to the AD process, research data shows that there is a relationship between the decrease of testosterone levels in serum and the clinical diagnosis of AD, indicating that testosterone levels may be a significant part in the process of AD [70]. The decreasing testosterone level is closely related to AD in elderly men. Research on 5,251 elderly healthy men and 240 elderly men with AD disclosed low testosterone levels in serum were significantly associated with increased risk of AD in elderly men (p = 0.006) [71]. All this evidence further confirmed the correlation between testosterone levels and AD.
AVPR1A gene, together with AVPR2, was found to have a relatively significant relationship with neuroprotection according to computational and biological analysis. Both AVPR1A and AVPR2 are the targets of the pharmacologically tested drug conivaptan. Actually, conivaptan is a non-peptide small molecular antagonist of vasopressin receptor, inhibiting the two isotypes of the vasopressin receptor, V1a and V2. It was approved in 2004 by FDA and used as its hydrochloride salt for the treatment of hyponatremia (low blood sodium levels) caused by syndrome of inappropriate antidiuretic hormone, SIADH [72]. The CvR value of conivaptan in the pharmacological experiment results reached 1.2465, indicates that conivaptan has significant neuroprotective effects potentially produced by two targets, AVPR1A and AVPR2. Our results revealed that AVPR1A and AVPR2 could be druggable targets for AD and exert neuroprotective effects under the antagonism of conivaptan. Therefore, conivaptan could be a key drug in the development of anti-AD drugs. In the computational simulation study for anti-AD drugs repurposing, a virtual screening experiment for 1,553 drugs found that conivaptan has a higher docking score among the docking scores of 1,553 drugs [73]. This demonstrated that conivaptan has great potential as a novel drug for anti-AD drug research. Simultaneously, relevant experimental studies have shown that conivaptan can not only reduce brain edema and the damage of the blood-brain barrier, but also improve the neurological functions deficits after ischemia in mice [74]. Therefore, it is very feasible to repurpose conivaptan in the drug discovery process of AD.
Notably, researchers from the Gladstone Institute screened more than 1,300 FDA approved drugs recently and this new experimental and computational drug repurposing study found that bumetanide can significantly slow down the generation of AD symptoms in AD mice models, and clinical trials of bumetanide for the treatment of patients with AD will be conducted before long [75]. Bumetanide is a loop diuretic regulating the ability of cells to absorb sodium ions and chloride ions, it is similar to the pharmacological effects of conivaptan. Bumetanide and conivaptan are both closely related to antidiuretic hormone, implying that conivaptan could also have that potential to be an anti-AD drug and its targets AVPR1A and AVPR2 can be essential roles in the process of AD.
Although these drugs discussed above are potentially effective against oxidative stress and AD, the doses should be carefully treated for drug repurposing. In this study, therapeutic effects of these experimental drugs were based on the SH-SY5Y cell model, and the effective dose in vitro might not be converted to dose in vivo directly. If these drugs are tested in animal models of AD or even in humans, the doses that achieve therapeutic effects will be different. Therefore, when the dose of a drug to achieve a therapeutic effect is higher than the recommended dose, we need to consider not only the efficacy of the drug, but also the safety concerns. These issues need to be further verified by animal experiments.
Advantages and perspectives
In this study, we identified several potential genes related to neuroprotection and AD based on phenotypic screening data and polypharmacology networks. From the perspective of oxidative stress hypothesis of AD, we then employed a systematic workflow to uncover some potential targets associated with neuroprotective effects. Besides, several drugs were also found to exert potential neuroprotective effects against oxidative stress, even AD. Phenotypic screening data based active compounds discovering method had been used in previous studies. For example, a two-stage phenotype-based screening research in a zebrafish model of Dravet syndrome successfully identified synthetic cannabinoid compounds with anti-seizure activity [76]. However, this research did not take pathogenic mechanisms and therapeutic targets into consideration. A phenotypic screening experiment on zebrafish was conducted to screen new antitumor metallodrugs with anti-cell proliferation activity [77], but there was also no further analysis of the mechanism against proliferation. In our study, on the basis of phenotypic screening data on oxidative stress neurons, our computational methods further predicted the pathogenic mechanisms and therapeutic targets for AD. Additionally, our predictions were based on real-world phenotypic screening data obtained from pharmacological experiments. Most importantly, our method could also be applied in other complex multi-mechanism diseases with broad application prospects.
There are some limitations in our research as well. For example, the neuroprotective effects and correlation with AD of these genes outside the DTI networks cannot be discovered yet, and computational prediction accuracy also has room for further improvement if more phenotypic screening data can be obtained.
Nevertheless, we are now actively developing novel methods that could predict therapeutic targets outside the DTI networks by integrating multi-scale biomedical data such as other types of phenotypic screening data and gene expression profiles.
Conclusions
In this study, we proposed a workflow to integrate phenotypic screening data and polypharmacology networks, in combination with computational and bioinformatical analysis. Some potential therapeutic targets for AD, such as NR3C1, SHBG, ESR1, PGR, and AVPR1A with significant neuroprotective effects were identified from the perspective of oxidative stress. Our predictive results especially showed that NR3C1, as a neuroinflammation-related gene, has great potential to be part of pathogenic mechanisms and a therapeutic target for AD. Drugs or drug-like compounds with neuroprotection were also found by conducting pharmacological experiments, such as beclometasone, methylprednisolone, and conivaptan. We further analyzed the targets and drugs with neuroprotective effects, and enriched biological pathways for identifying pathways associated with AD. In addition, our methods used in this study can help uncover multiple targets with neuroprotective effects simultaneously and discover some of the most potential effective therapeutic targets for AD. Based on the real-world phenotypic screening data obtained from in vitro pharmacological experiments, our computational predictions and analysis can help identify some novel and promising therapeutic targets. And these potential neuroprotective targets we identified in this study may exert significant effects in the process of anti-AD drug discovery. Our systematic workflow combining pharmacological experiments and computational methods can also provide useful applications to many other complex diseases.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
This study was supported by the National Natural Science Foundation of China (Grants 82173746 and 82104066), Shanghai Frontiers Science Center of Optogenetic Techniques for Cell Metabolism (Shanghai Municipal Education Commission, Grant 2021 Sci & Tech 03–28) and the 111 Project (Grant BP0719034).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
Data that support the findings of this study are available in the supplementary materials.