
Abstract
Background:
Advanced age is the greatest risk factor for the development of Alzheimer’s disease (AD). This implies that some aspect of the aged milieu is possibly accelerating the development of AD related pathologies.
Objective:
We hypothesized that intracranially injected with AAV9 tauP301L may cause a greater degree of pathology in old versus young mice.
Methods:
Animals were injected with viral vectors overexpressing the mutant tauP301L or control protein (green fluorescent protein, GFP) into the brains of mature, middle-aged, and old C57BL/6Nia mice. The tauopathy phenotype was monitored four months after injection using behavioral, histological, and neurochemical measures.
Results:
Phosphorylated-tau immunostaining (AT8) or Gallyas staining of aggregated tau increased with age, but other measures of tau accumulation were not significantly affected. Overall, AAV-tau injected mice had impaired radial arm water maze performance, increased microglial activation, and showed evidence of hippocampal atrophy. Aging impaired open field and rotarod performance in both AAV-tau and control mice. The efficiency of viral transduction and gene expression were the same at all animal ages.
Conclusion:
We conclude that tauP301L over expression results in a tauopathy phenotype with memory impairment and accumulation of aggregated tau. However, the effects of aging on this phenotype are modest and not detected by some markers of tau accumulation, similar to prior work on this topic. Thus, although age does influence the development of tauopathy, it is likely that other factors, such as ability to compensate for tau pathology, are more responsible for the increased risk of AD with advanced age.
INTRODUCTION
In Alzheimer’s disease (AD), neurofibrillary tau tangles (NFTs) are one of the cardinal pathologies associated with neuronal loss and declines in cognition [1 –4]. The risk of developing symptoms of AD increases as individuals age [5]. This occurs concurrently with systemic aging, including basal increases in innate immune activation, sometimes referred to as “inflammaging” [6, 7]. Immune activation is related to neurodegenerative diseases such as AD [8, 9] and may accelerate tauopathy [10, 11]. Genome wide association studies have validated a crucial link between innate immune marker polymorphisms and the risk of AD [12, 13]. Therefore, making the importance of assessing the role of age-associated physiological changes, including immune backgrounds, when examining the development of tau pathology paramount.
Germ line transgenic models of tauopathy have been of considerable value in understanding the stages of tau accumulation in brain tissue, but these typically express mutations that produce pathological changes at a relatively fixed age, often early in the life course. Most transgenic models do not allow age of onset to be an independent variable in experiments. The use of adeno-associated viral vectors (AAV) to introduce transgenes overcomes this limitation by initiating expression at any age. Several studies have demonstrated that this intracranial AAV methodology is successful at producing tauopathy and neurodegeneration. Studies have employed both mutant and wild-type tau to promote tauopathy. Some of the earlier studies first demonstrated the utility of AAV induced tauopathy models in rats and reported tau conformational changes [14], neuronal loss [14 –16], behavioral deficits [14 –16], and glial activation [14]. Other studies injected AAV tau constructs into mice and similarly reported substantial neuron loss, formation of NFTs, cortical atrophy, gliosis, and synapse loss [17 –20]. These pathological features were accompanied by behavioral changes that showed abnormalities in open field, y-maze, and fear conditioning demonstrating increased anxiety, and a reduction in learning and memory [21 –23]. Additionally, AAV tau vectors have been used to study propagation of tauopathy across synapses [20–22 , 25].
The purpose of this study was to invesigate the influence of chronological age of pathology onset on behavior, histology and neurochemistry to evaluate the tauopathy phenotype in a viral mouse model. We also evaluated the possible effects of age on AAV transduction efficiency and transgene expression. Ages were selected that represented mature (8 months), middle-aged (12 months) and old (16 months) mice to avoid influences that may be more related to development or maturation in which to study AAV9 tauP301L expression.
MATERIALS AND METHODS
Animals and experimental design
Male and female C57/BL6 mice (n = 69) were ordered from the National Institute of Aging colony. All animals were kept on 12-h light and dark cycles and were provided food and water ad libitum. Three different age groups were compared: mature (aged 8 months, n = 20), middle-aged (aged 12 months, n = 20), and old (aged 16 months, n = 29). The increased sample size for the old group was to maintain statistical power due to anticipated attrition over the duration of the study. Half of each age group received an experimental injection of AAV9 tauP301L and the remainder received a control injection of AAV9 green fluorescent protein (GFP). Supplementary Figure 1 provides the experimental flowchart and sample sizes for each measurement with the total number of female and male mice in each group. Animals were monitored weekly for signs or symptoms of declining health such as weight loss for the duration of the study. Three and half months after surgery, behavioral assessments were conducted which included open field, rotarod, and the radial arm water maze (RAWM). Two weeks after behavioral assessments, animals were euthanized, and tissues were collected. One hemisphere was dissected and processed for neurochemical measurements and the other hemisphere was prepared for histological measurements of tauopathy and other pathological markers.
Viral preparation and stereotaxic injections
A glycerol stock of competent SURE2 cells containing GFP plasmid was donated by Dr. Kevin Nash from the University of South Florida. Then 2N4R tauP301L gene fragment was ordered from Genewiz (South Plainfield, NJ, USA). Both viral constructs contained cytomegalovirus enhancer, chicken β actin promoter, woodchuck hepatitis virus posttranscriptional regulatory element, and the experimental condition gene (tauP301L or GFP) [26, 27]. The plasmids’ DNA were sequenced and confirmed to be correct. HEK293T cells were co-transfected with adenovirus helper elements, and a plasmid that encodes the AAV9 capsid proteins. Viral purification was completed with iodixanol gradients. The viral vectors were concentrated using Amicon Ultra-15 centrifugal filters (Sigma, Burlington, MA, USA; Cat. No. UFC910024) in Dulbecco’s phosphate buffered saline (DPBS; pH 7.4). Both the GFP and tauP301L constructs were titered using RT qPCR with primers targeted to the woodchuck hepatitis virus posttranscriptional regulatory element sequences. The viruses were diluted to 8.8×1011 vector genomes (vg)/μL with DPBS. The total volume was divided into 10μL aliquots and stored at –80°C. For each day of surgery, a new frozen aliquot was removed to minimize freeze thaw cycles. To detect the region transduced with AAV by the injection of AAV9, 8.8×1010 vg/ml (a 1/10th dosage) of AAV GFP was added to tauP301L viral vector solution.
At 8-, 12-, or 16-months of age, animals received bilateral stereotaxic injections of AAV to the anterior portion of the cortex (coordinates ML: ±2.20, AP:2.2, DV: –3.0 relative to bregma) and hippocampus (coordinates ML: ±2.7, AP: –2.7, DV: –3.0, relative to bregma). Two μL of viral solution were administered at each site at a rate of 0.5μL per minute using a stereotaxic mounted pump. Injections were administered by a glass pulled pipette needle attached to a 10μL Hamilton syringe (Hamilton, Reno, NV; cat. no. 80000) with heat shrink tubing. All animals were placed on a heating pad to maintain body heat and observed during postoperative recovery.
Behavioral assessments
At 3.5 months of viral incubation, behavioral assessments were conducted. All assessments were performed by an experimenter blinded to age and injection group. There were 5 animals that died prior to the behavioral assessments; these were from the mature GFP (n = 1), old GFP (n = 1), and old tauP301L (n = 3) injection groups. All mice were weighed and monitored weekly to acclimate animals for routine behavioral handling. The open field task was conducted first. Animals were placed into an open field enclosure (L 40 mm/ W 40 mm/H 40 mm, BioSeb; Vitrolles, France) for 10 min and the total distance traveled was measured by AnyMaze software (Stoelting, Wood Dale, IL, USA; cat. no. 60000). The second task was the rotarod, where animals are placed on a TSE rotating rod apparatus (TSE Systems, Chesterfield, MO, USA) that increased in speed (from 4 to 40 rpm) over 5 min to evaluate motor performance. All animals completed 4 trials per day over 2 days. There was a minimum of 20 min rest between trials to avoid muscle fatigue. Fall latency was recorded and averaged across trails for analysis.
The final behavioral test was the RAWM that was used to evaluate visuospatial navigation memory. This was conducted as described by Alamed et al. [28]. This test included four tasks over four days: training, testing, reversal, and open pool. The maze for this series of tasks was a 6-arm circular water maze. External visual cues were placed on the walls that allow the animals to orient themselves spatially. Additionally, the experimenter location also served as an extramaze cue during this task. The start arm changed for each trial and was varied between animals to minimize olfactory cues. The goal arm that contained the platform, remained the same for all the trials for each individual mouse, but was also moved between mice. Two platforms were used, a visible platform that sat above the water and a hidden platform that was just below the surface of the water. The platform served as a reward to allow the animal to escape the need to swim. On each day there were 15 trials that were each 1 min in duration for the animal to find, swim to, and ascend the platform, with intertrial and interblock intervals designed to minimize risk of fatigue (as in [28]). On the training day, the visible and hidden platforms were alternated in the goal arm. Only on the training day, if the animal was unable to reach the platform within 60 s, they were gently guided to the platform by the experimenter. An arm entry was defined as all four limbs within the maze arm and incorrect arm entries were documented for each trial. The second day was the testing task where only the hidden platform was used for 15 trials as described above for the training task above. The reversal task was administered on day 3, where the goal arm was now moved to the arm 180° from the original goal arm. Again, only the hidden platform was used, and incorrect arm entries were noted, over 15 trials. On the final task, to assess the animals’ ability to swim, see, and ascend the platform, an “open pool” was used. The visible platform was placed in the center of the maze and the external cues were removed. The latency to ascend the platform was recorded. For the first 3 tasks, the total number of errors were used for analyses. For the open pool task, the latency to ascend the platform was used for analysis.
There was a total of 25 animals excluded from the RAWM task using predefined exclusion criteria. Six mice had visible eye abnormalities consistent with blindness and were not tested. We previously demonstrated blind mice are unable to learn the task [28]. Twelve were not able to swim adequately on the training day and testing were excluded. Four had overt seizures on the days of testing and were discontinued. Finally, on the last day of testing, three mice were unable to reach and ascend the open pool visible platform within 20 s on the final three trials and were deemed to have performance deficits. It is important to note that the exclusions were in both the old GFP and old tauP301L injection groups. Half of the individuals in those groups were excluded and the most common cause was inadequate ability to swim (60%). After applying the exclusion criteria, the final sample sizes for RAWM were mature GFP n = 9, middle-aged GFP n = 7, old GFP n = 5, mature tauP301L n = 10, middle-aged tauP301L n = 5, and old tauP301L n = 6 (see Supplementary Figure 1). It should be noted that the proportion of mice requiring exclusion was greater in these inbred C57BL/6 mice than in our prior work at these ages using APP+PS1 mice on a mixed genetic background [29].
Tissue collection and sample preparation
Tissues were collected two weeks after the behavioral assessments. Mice were overdosed with diluted Euthasol® solution (3.9 mg/kg of pentobarbital and 0.5 mg/kg of phenytoin). Once mice were completely anesthetized, they were transcardially perfused with 25 mL 0.9% normal saline while on a heating pad to maintain body temperature to reduce artefactual tau phosphorylation due to hypothermia [30]. The brains were immediately extracted and placed on an iced stage and bisected sagittally. The left hemispheres were immersed in 20 mL of freshly prepared phosphate buffered 4% paraformaldehyde (pH 7.4) for 24 h at 4°C. The hippocampus and anterior aspect of the cortex of the right hemispheres were microdissected and frozen on dry ice. Samples were stored at –80°C until samples were homogenized. After fixation, the left hemispheres were cryoprotected by serial immersion in increasing sucrose concentrations (10%, 20%, and 30%) for 24 h each. The fixed hemispheres were kept at 4°C until sectioned. For sectioning, the hemispheres were placed on a freezing microtome and sectioned horizontally at a 25μm thickness. The only exception to this was every 10th section was 50μm thickness to allow for volumetric analyses. All sections were stored at 4°C in DPBS with 10 mM sodium azide solution (7.4 pH) until stained.
For ELISA assays, the microdissected hippocampi were homogenized in 10 volume/weight of RIPA homogenization buffer (50 mM Tris-Cl, 150 nM NaCl, 1% NP40, 0.1% SDS, 0.05% sodium deoxycholate) that contained protease inhibitor cocktail (Sigma Aldrich, St. Louis, MO, USA; Cat. No. P8340), deacetylase inhibitor cocktail (MedChem Express, Monmouth Junction, NJ, USA; Cat. No. HY-K0030) phenylmethanesulfonyl fluoride (Sigma Aldrich, St. Louis, MO, USA; Cat. No. P7626), and phosphate inhibitor cocktails II and III (Sigma Aldrich, St. Louis, MO, USA; Cat. Nos. P5726 & P0044). Samples were first homogenized using a handheld mechanical rotating pestle homogenizer and then sonicated. A 25μL aliquot of the crude homogenate was centrifuged at 50,000 g at 4°C for 1 h. The supernatant was collected and stored at –80°C as the detergent soluble fraction. The detergent insoluble pellet was further processed by resuspending in 70% formic acid with the handheld mechanical rotating pestle. Samples were incubated on a tube revolver for 30 min, and neutralized with 1 M tris-base (pH 7.6). Each sample was individually adjusted to a pH of 7 with 50% NaOH. The detergent insoluble samples were stored at –80°C. Pierce BCA protein assays were used to determine the protein concentrations for ELISA assays.
ELISA assays
The detergent soluble and detergent insoluble fractions were used for measurement of total tau (Fisher Scientific, Cat. No. KHB0041), phosphorylated tau Ser199 (Fisher Scientific, Cat. No. KHB7041), and phosphorylated tau Ser396 (Fisher Scientific, Cat. No. KHB7031). The detergent soluble fraction was used to quantify GFP (Abcam Cat. No. Ab171581). Samples were first diluted 1000x in RIPA buffer and then further diluted in the buffer provided by each kit to the empirically determined concentration to be on the linear portion of the standard curve. An additional sandwich ELISA for soluble oligomeric tau (using TOC1 antibody) was performed for the detergent soluble fraction [31]. For the TOC1 ELISA, samples were diluted 25x. All ELISA concentrations were normalized to the total protein from the Pierce BCA protein assays.
RNA measurement by reverse transcriptase quantitative polymerase chain reaction
Total RNA was isolated from each anterior cerebral cortex microdissection using the Qiagen RNeasy mini kit (Germantown MD) according to the manufacturer’s directions. Briefly, 10–30 mg wet weight was homogenized in 600μl of the provided guanidine-thiocyanate buffer containing 143 mM beta-mercaptoethanol using a motorized pestle fitting into a microfuge tube. Debris was removed by centrifugation (10,000× g for 3 min). The supernatant was applied to mini spin columns which enrich for RNA >200 bases, washed several times with ethanol, and eluted in 50μl RNAse-free water. Concentration was assessed using a Nanodrop One spectrophotometer. For each sample, 100 ng of total RNA was reversed transcribed using the BioRad iScript cDNA kit as outlined by the manufacturer. The kit mastermix includes a modified Moloney murine leukemia virus reverse transcriptase, RNase inhibitor, oligo (dT), and random hexamers. Applied Biosystems TaqMan gene expression assays were purchased from ThermoFisher (Waltham MA; MAPT; FAM; Hs00213491_m1; GAPDH, VIC, primer limited; Mm99999915_g1). DNA polymerase was activated at 95°C for 3 minutes, and PCR conditions included 40 cycles of 95°C for 15 s, then 60°C for 30 s. Samples that failed to amplify by 40 cycles were assigned a Ct value of 40. The delta-delta Ct method was used to compare relative expression in each treatment condition, and values were normalized to the mature GFP group.
Histopathology
For immunohistochemical stains, sections were placed into free-floating section staining racks and conducted as previously described by Gordon et al. [32]. The sections were first blocked to eliminate endogenous peroxidase (10% methanol, 3% H2O2 in phosphate buffered saline pH 7.4; PBS) and washed with PBS. Next, sections were permeabilized (0.2% lysine, 1% Trition X100, 4% normal goat serum in PBS) and placed into primary antibody (total tau, HT7, Invitrogen MN1000B, 1:5k; phosphorylated tau Ser202/Thr205 AT8, Invitrogen MN1020B, 1:10k; phosphorylated tau Ser396, Anaspec AS 54977-025, 1:30k; MHCII, BD Pharmingen 556999, 1:5k; GFP Abcam ab13970 1:30k). Incubations were performed overnight at room temperature. The following day sections were removed from the primary antibody and washed with PBS. Then sections were placed in secondary antibody (goat anti-rabbit IgG, Vector Laboratories BA-1000; goat anti-rat IgG, Vector Laboratories BA-9400; goat anti-chicken IgY, Vector Laboratories BA-9010) for stains where the primary antibodies were not biotinylated for 1 h and then washed with PBS. Sections were then placed into a Vectastain® Elite® ABC kit solution (Vector Laboratories, Burlingame, CA; cat. no. PK-6100). Sections were washed with PBS and tris buffered saline pH 7.4. Then, sections were placed into a color development solution (0.05% diaminobenzidine, 0.5% Ni2+, 0.03% H2O2 in TBS) for up to 5 min. To end the development, sections were removed and washed with tris buffered saline (pH 7.4) and PBS again. Sections were then mounted on positively charged microscope slides (Fisher Scientific, Waltham, MA; cat. no. 1255015), dehydrated, and coverslipped with DPX mounting medium (Electron Microscopy Sciences, Hatfield, PA; cat. no. 13512).
Both cresyl violent and Gallyas silver stains were conducted on sections previously mounted on slides that were dried overnight. Cresyl violet stains the ribosomal RNA of the Nissl bodies and was conducted on the 50μm slides for hippocampal volumetric analysis. The day of cresyl violet protocol, slides were briefly rehydrated. Slides were immersed in a 0.05% cresyl violet dye and destained with acetic acid water solution (pH 3.5) to remove excess background staining. Slides were washed with water and then immediately dehydrated and coverslipped.
Gallyas silver stain targets the silver positive NFTs [33]. After briefly rehydrating slides, slides were placed in 5% periodic acid for 5 min and washed with water. Then tissues were placed in a silver enhancing solution (10% potassium iodide, 4% NaOH, 0.035% silver nitrate) for 1 min. Next, tissues were placed in 0.5% acetic acid solution for 10 min. Slides were immersed in the working developer solution (2.5% sodium carbonate, 0.1% ammonium nitrate, 0.1% silver nitrate, 0.5% tungstosilic acid, 0.2% formaldehyde) for 10 min and upon development, slides were washed with acetic acid and water. Then slides were placed in a gold toning solution (0.1% gold chloride solution) and washed with water. Finally, tissues were incubated with 0.1% sodium thiosulphate and washed with water. Slides were then dehydrated and coverslipped.
Once the mounting medium was dried for all slides, they were scanned at 200x magnification with the ZEISS Axio Scan.Z1. The entire tissue section was analyzed using the NearCYTE image analysis software (NearCYTE, Pittsburgh, PA). Analysis was conducted by first creating a segmentation file that identified positively stained pixels selecting hue-saturation-intensity parameters in the full range of staining intensities for the experiment. The ratio of positive staining to the total tissue section was calculated (percentage area) and was used for analyses. For the volumetric analysis of the cresyl violet staining, the Cavaleri equation was employed, V = A×T×t, where A = sum of the hippocampal areas (μm2) on the sections analyzed, T = section interval (number of sections), and t = section thickness (50μm).
Statistical analyses
Statistical analyses were conducted with IBM SPSS Statistics 25 (Armonk, NY, USA) and graphical figures were made with GraphPad Prism 9 (La Jolla, CA). For all tau biochemical and histological measurements, one-way ANOVAs were conducted to determine if there were any age effects between mature, middle-aged, and old tauP301L injected groups. The additional markers of pathology that included MHCII staining, hippocampal volume, and hippocampal weight were included for all three age groups for both GFP and tauP301L injected groups; two-way ANOVAs were conducted to identify any main effects of age and or injection. For open field and rotarod, two-way ANOVAs were conducted to determine if there were significant main effects of injection and/or age. For the first three RAWM tasks (training, testing, and reversal), the errors were collapsed for the total completed errors for each individual day and two-way ANOVAs were conducted. For the open pool RAWM task, a two-way repeated measures ANOVA was conducted. All significant F tests were followed with Fisher’s Least Significant Differences post hoc analyses. Unpaired t tests were conducted to determine sex differences for all outcome measures. Pearson’s correlations (r) were conducted to determine if there were any significant correlations between histological tau measurements with hippocampal volume and immune activation (MHCII).
RESULTS
Behavioral measures
In addition to routine handling, we acclimated mice to behavioral testing paradigms by first evaluating performance in the open field. There was a main effect of age (F(2, 58) = 5.46, p < 0.01), but not injection group. Post hoc comparisons indicated the old animals travelled significantly less distance (mean [M] = 29.67, standard error of the mean [SEM] = 2.941) when compared to the mature (M = 40.70, SEM = 3.377; p < 0.05) and middle-aged (M = 43.17; SEM = 2.943; p < 0.01) animals (Fig. 1A). Next, motor performance was measured by the averaged fall latency on the rotarod task. There was a main effect of age (F(2,58) = 13.75, p < 0.0001) but not injection group. With further examination, it was evident that the AAV GFP injected animals had an age-dependent decline in motor performance, where the mature GFP animals had the greatest capacity to remain on the rod (M = 201.3, SEM = 16.37) compared to the old GFP group (M = 75.88, SEM = 16.62) (Fig. 1B). Note that some of the old group mice had very little capacity to remain on the rotating rod with latencies close to zero. When age differences were examined exclusively in the AAV tauP301L injected animals, there were significant differences (F(2,29)=3.354, p = 0.049), limited to the mature (M = 158.3, SEM = 20.31) and old (M = 57.02, SEM = 17.30) animal comparison (p = 0.023).

Effects of AAV tauP301L and AAV GFP on open field and rotarod performance. A) There were significant age-related reductions in total distance traveled in both the AAV GFP and AAV tauP301L injection groups for the open field task. B) Similarly, there were significant age-related reductions in time to fall for both the AAV GFP and AAV tauP301L injection groups on the rotarod task. Data are presented as mean±SEM (error bars). Each dot represents one mouse. * p < 0.05, ** p < 0.01, *** p < 0.001 by Fisher’s Least Significant Differences post hoc analysis.
Visuospatial navigation memory was assessed by the number of incorrect arm entries (errors) on the RAWM tasks. Because of the potential health risk the swimming tasks pose for the older mice, mice with certain performance deficits (see exclusion criteria in Methods) were not tested or removed during testing and completely excluded from data analysis. This resulted in reduced samples sizes primarily for both old age groups (Supplementary Figure 1). On the day 1 training task, there were no significant differences in errors for either injection group (F(1,33) = 0.024, p = 0.88) or age (F(2,33) = 0.225, p = 0.88; Supplementary Figure 2). In contrast, on the day 2 testing task (Supplementary Figure 3), there was a significant overall effect of age (F(2,33) = 7.38, p < 0.01) and interaction of age and injection group (F(2,33) = 3.61, p = 0.038). However, upon post hoc analyses, these age effects were restricted to the AAV GFP injected mice. Both the old AAV GFP mice and the old AAV tau mice performed at chance levels (50 errors using data from blind mice, [28]), and were not significantly different. Because of the “ceiling” effects and small sample sizes in the oldest age group, we conducted a two-way ANOVA excluding the old AAV GFP and AAV tau groups (Fig. 2). There were significant main effects of age (F(1,27) = 4.60, p = 0.01) and injection group (F(1,27) = 12.84, p = 0.001) with the AAV tau mice performing more poorly than the AAV GFP injected in both the mature and middle aged groups. The posthoc means analysis confirmed the effects of injection group at each age, but no effects of age within the AAV GFP group or the AAV tau group were significant. On day 3 a reversal task was conducted, which involves relocating the platform 180 degrees from the prior location. There were no significant differences between the injection groups (F(1,36) = 2.260, p = 0.14) or age (F(2,36) = 0.69, p = 0.50; Supplementary Figure 4). On day 4, the time to swim to, and ascend the visible platform was analyzed. There were no significant main effects of injection (F(1,33) = 0.200, p = 0.66) or age (F(2,33) = 0.022, p = 0.98) on this estimate of task performance capability for the mice remaining in the analysis (Supplementary Figure 5). We found no effects of sex on any of the RAWM results. We also found that similar numbers of male and female mice were removed from the analysis due to the exclusionary criteria described in methods (p > 0.17).
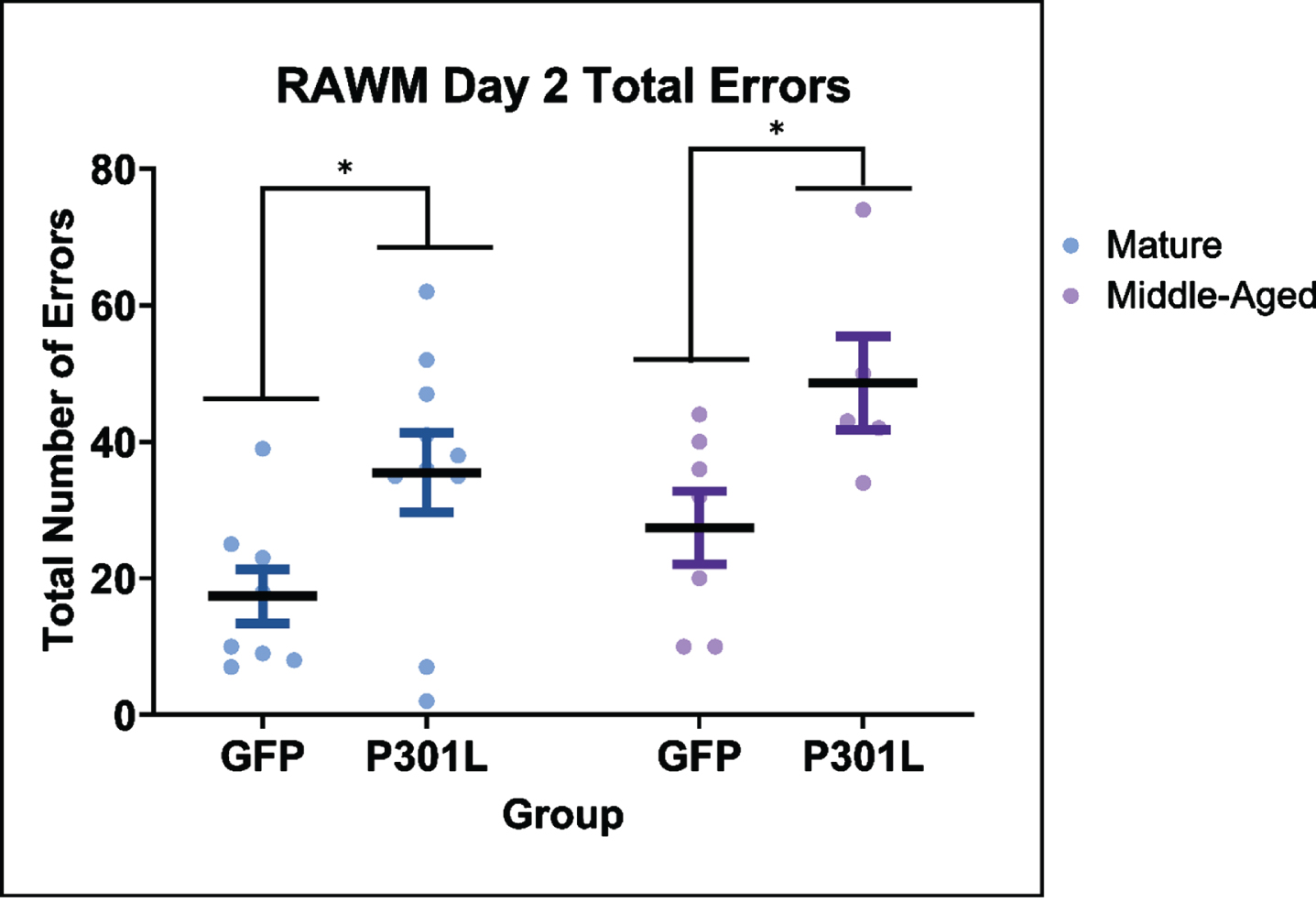
Effect of virus injection in mice of different ages on radial arm water maze (RAWM) performance in the testing task on day two. The mature and middle-aged AAV tauP310L mice made more errors than the mature and middle-aged AAV GFP mice. Data are presented as mean (symbol)±SEM (error bars). * p < 0.05 Fisher’s Least Significant Differences post hoc analysis.
Histology
To estimate if age influenced tau accumulation, tissue sections were stained for several tau markers, including total human tau (HT7), phosphorylated tau Ser202/Thr205 (AT8), phosphorylated tau Ser396, and silver stains according to the Gallyas method (Fig. 3). The sections were digitally imaged and analyzed for the area of staining. Total tau immunostaining and phosphorylated tau Ser396 immunostained areas were similar at all three ages (p = 0.60; Fig. 4A; p = 0.44; Fig. 4C respectively) of the tau mice. However, the phosphorylated tau Ser202/Thr205 (antibody AT8) staining was significantly affected by mouse age (F(2,28) = 3.13, p < 0.05), with the old animals (M = 0.45, SEM = 0.008) having a greater amount of staining when compared to the mature animals (M = 0.25, SEM = 0.003; Fig. 4B). To further examine pathological tau, we measured silver positive fibrils by Gallyas staining. There was a significant overall ANOVA effect of age, (F(2,29) = 7.170, p < 0.01) (Fig. 4D) and post hoc tests found the old age group had significantly greater Gallyas staining than the mature (p < 0.01) and middle-aged (p < 0.01) groups.

Histological staining for tau markers. Mouse brain sections were stained for HT7 (human total tau; A-C), phosphorylated tau Ser202/205 (AT8; D-F), phosphorylated tau Ser396 (G-I), and by the Gallyas silver method for NFT (J-L). Each panel shows a single representative horizontal section with an inserted higher magnification of the hippocampus. The left column (A, D, G, J) represents specimens from mature mice, the middle column (B, E, H, K) includes sections from middle-aged mice, while the right column depicts aged mice (C, F, I, L). Scale bar in J represents 1000μm for low magnification and 300μm for inserts.

AT8 labeled tau and Gallyas staining increase with age, but HT7 and phosphorylated tau Ser396 immunostaining do not. Scatterplots of individual animal’s expression values after injection of AAVtaup301L are plotted by age group. Quantification of positive fractional area for (A) total tau (HT7) staining, (B) phosphorylated tau Ser202/Thr205 (AT8) staining, (C) phosphorylated tau Ser396 staining, and (D) silver positive Gallyas neurofibrillary tau tangle staining. Data are presented as mean±SEM (error bars). For each marker, values from 8 sections per mouse were averaged and plotted so that each dot represents one mouse. * p < 0.05, ** p < 0.01 Fisher’s Least Significant Differences post hoc analysis.
Immune activation was measured histologically by MHCII staining and demonstrated significant main effects of both injection group (F(1,58) = 13.76, p < 0.01) and age (F(2,58) = 4.76, p < 0.05; Fig. 5A). Overall, the AAV tau injected mice (M = 0.041, SEM = 0.005) had twice the amount of MHCII staining when compared to the AAV GFP injected mice (M = 0.020, SEM = 0.005). In addition, there were effects of age within both injection groups with the mature animals (M = 0.020, SEM = 0.005) had significantly less staining compared to middle-aged (M = 0.036, SEM = 0.005; p< = 0.05) and old (M = 0.039, SEM = 0.004; p < 0.01) animals. Although both AAV-GFP and AAV-tauP301L groups displayed age-related changes, the AAV tauP301L injected animals did so to a greater extent. To estimate atrophy associated with the AAV tau injections, hippocampal volume was measured using Nissl stained sections (F(1,58) = 18.178, p < 0.01; Fig. 5B) and dissected hippocampal weights (F(1,58) = 18.96, p < 0.01; Fig. 5C). Both measures indicated significant injection group effects, where the AAV tauP301L injected groups had reduced volumes compared to the AAV GFP injected animals. However, there were no significant effects of age in either injection group.

Immune response and hippocampal atrophy after AAV GFP or AAV tauP301L. A) Quantification of MHCII staining demonstrated an age dependent increase in positive staining and an injection group difference. B) Quantification of hippocampal volume estimated using 8 sections through the entire region by the method of Cavalieri indicated that the tauP301L mice had a significant reduction of hippocampal volume compared to the AAV GFP injection group. C) The weights of dissected hippocampi measured in milligrams demonstrated that AAV tauP301L had a significant reduction in hippocampal weight when compared to the AAV GFP group. Data are presented as mean±SEM (error bars). Each dot represents one mouse. * p < 0.05, *** p < 0.001 Fisher’s Least Significant Differences post-hoc analysis.
When sex differences were examined, the significant differences were limited to histological measures of total tau (HT7) and silver NFTs (Gallyas) (Supplementary Figure 5). Across the three age groups female tauP301L injected mice had a greater amount of total tau staining (Supplementary Figure 5A) and silver NFTs (Supplementary Figure 5B) compared to males (ps < 0.012). When examining the specific age groups, only the old female tauP301L injected mice had greater Gallyas staining compared to the male mice (p = 0.012). Furthermore when examining if there were significant correlations in the tau measurements of HT7 and Gallyas staining with hippocampal volume (r = –0.42, p = 0.0003 and r = –0.4801, p < 0.001 respectively) and histological MHCII (r = 0.41, p = 0.0006 and r = 0.31, p = 0.012 respectively).
One potential artifact we wished to exclude was an age-associated change in the transduction volume of the injected AAV viruses. We estimated this parameter by measuring GFP immunostaining, with the rationale that, in contrast with AAV tau, GFP would not aggregate, propagate, or accumulate over time. For the AAV tauP301L injected mice we added a 1/10th dose of AAV GFP to evaluate the extent of the injection area. As expected, the full dose AAV GFP control injected animals had a significantly greater intensity of staining when compared to the AAV tauP301L injected animals (F(1,58) = 495.21, p < 0.01; Fig. 6). However, there was no effect of age on the extent of GFP staining in either group. The data also resulted in an average coefficient of variation of 13% (standard deviation (SD)/M; average of all groups) indicating relatively consistent injections of the AAV constructs.

Quantification of positive fractional area for GFP staining. The AAV GFP injected animals had greater GFP staining when compared to the AAV tauP301L injected animals with no effect of age. Scatterplots of individual animal’s expression values are plotted by injection and age groups. Data are presented as mean±SEM (error bars). Each dot represents one mouse. *** p < 0.00001 Fisher’s Least Significant Differences post hoc analysis.
Neurochemistry
We conducted RT qPCR for human MAPT RNA to determine whether age had an impact of the efficiency of tau transgene expression, which might account for increases observed histologically. However, the anterior cortex RNA demonstrated that there were no significant differences with age for the AAV tauP301L injected animals (F(2,29) = 0.304, p = 0.74; Fig. 7).

There was no effect of age on the human MAPT RNA levels in the AAV tauP301L injected animals. RNA expression was normalized to the mature control animals. Data are presented as mean±SEM (error bars). Each dot represents one mouse. Fisher’s Least Significant Differences post hoc analysis.
When hippocampal tau markers were examined by ELISA, there were no significant age-effects in the soluble total tau, and phosphorylated tau assays or the soluble oligomeric tau assay, using the antibody TOC-1 (Fig. 8A–D). Furthermore, there were no significant age differences in the insoluble tau markers of total tau (p = 0.84), phosphorylated tau Ser199 (p = 0.45), or phosphorylated tau Ser396 (p = 0.078; Fig. 9).

There was no effect of age in the AAV tauP301L injected animals for any tau marker in the detergent soluble cellular fraction. Tau concentration measured by ELISA assays in the detergent soluble hippocampal fraction normalized to total protein for (A) total tau using detection antibody recognizing both non-phosphorylated and phosphorylated epitopes (HT7), (B) phosphorylated tau Ser199, (C) phosphorylated tau Ser396, and (D) oligomeric tau recognized by the TOC1 antibody. Data are presented as mean±SEM (error bars). Each dot represents one mouse. Dashed line denotes the limit of detection.

There was no effect of age in the AAV tauP301L injected animals in any tau marker in the detergent insoluble cellular fraction, including (A) total tau, (B) phosphorylated Ser199, and (C) phosphorylated tau Ser396. Data are presented as mean±SEM (error bars). Each dot represents one mouse. Dashed line denotes the limit of detection.
DISCUSSION
The purpose of this study was to examine the effects of age upon the development of tau pathology when administering AAV9 tau P301L bilaterally into hippocampi and anterior cortices. The statistically significant findings demonstrated an increase of histological NFTs and phosphorylated tau (AT8) in the oldest age group compared to the middle-aged and mature mice. In addition, there was evidence of an age-dependent increase in immune activation that was even more pronounced in the tauP301L injected animals. Furthermore, the tau injected animals had reductions in hippocampal volume and hippocampal weight, but surprisingly there were no effects of aging. Other measures of tau were not significantly impacted by the host age. These included ELISA measures of soluble and insoluble tau (total, phosphorylated and oligomeric). There were also no significant changes in histological measures of total tau nor phosphorylated tau Ser396. There were main effects of aging on multiple behavioral assessments, yet these were generally similar in both injection groups. There were significant effects of age on locomotor performance (open field), motor control (rotarod), and spatial navigation (RAWM). However, when further examining spatial navigation performance in the RAWM, it was apparent that the AAVtauP301L injected mice performed worse than the GFP injected control mice at the mature and middle-ages. In the old mice, both treatment groups were performing close to chance resulting in a ceiling effect that was insensitive to any differences in spatial navigation capacity that might have existed in the two groups.
One possibility for increased tauopathy phenotype with age could result from reduced transduction efficiency of the AAV9 in aged animals, or reduced transgene expression. To examine these options, we tested for age effects on GFP expression in all mice, and for the human tau RNA expression in mice from the tauP301L treatment group. We found no differences with age in either measure, indicating, at least for these genes, the transduction efficiency was not reduced in aged mouse brain. This would imply that differences in the tau phenotype in older mice were associated with changes in the brain physiology responsible for promoting and preventing tau clearance or aggregation, rather than an artifact of AAV9 transduction efficiency with age. Nonetheless, these aging changes influencing the tau phenotype appear relatively modest.
Several other groups reported aging effects on tauopathy using viral vectors. Klein et al. [14] examined rats at 3 and 20 months and treated them with two doses in two brain regions for 2 weeks or 8 weeks. Although they concluded that there was a greater response to tau transduction in the aged rats, out of the 8 comparisons that included both ages, there was only one significant difference, and it was about 25% greater in the old rats. A second study by Wegmann et al. [24] injected AAV tauP301L into the entorhinal cortex and monitored spread into hippocampal neurons in 3-month-old versus 20-month-old mice. They reported greater spread to hippocampus in the aged mice, without increased tau expression in the entorhinal area (although there was more misfolding). Perhaps most related to this work is a recent publication in which some of us participated, Loon et al. [34]. This report compares responses to AAV expressing full length human wild-type tau or C-terminal truncated (aa 421) tau in mice treated from 4 to 8 months (young) with those treated from 12 to 16 months (middle-aged to old). They found increased histological Gallyas staining and AT8 staining with age, but no change in total tau immunostaining, consistent with the present manuscript. By western blot they found no change in total tau, phosphorylated tau Ser199/202 in either detergent soluble or insoluble fractions, again consistent with the present findings. In contrast, they also found increased phosphorylated tau Ser396 with age by both immunohistochemistry and western blot, unlike the present findings. They further observed neuron loss and impaired cognitive performance in the middle-aged/old group, but not the young group. In some instances, the changes were found only in the truncated tau group, but not the full length tau treatment.
One of the limitations of the current study is the inherent tissue injury that accompanies intracranial injections. The protocol requires a craniotomy, insertion of a needle, and the dispersion of injectate that all produce local tissue damage. Furthermore, the dispersion of the vectors creates a gradient of transduction around the injection site with higher transgene expression near the center and reduced expression at the periphery. The current study would have benefited from additional cognitive measures that did not require swimming abilities as approximately a third of the sample size demonstrated an inability to swim. Of those that were eliminated, most were old animals from both injection groups. This led to a reduction in sample size and could be contributing to the lack of significant injection group findings among the old animals for the RAWM. Finally, it would be beneficial to examine additional durations of transgene expression other than just four months of viral incubation. This may aid in identifying additional effects in biochemical and behavioral analyses.
In conclusion, these data validate the utility of AAV based transgenesis for the evaluation of the impacts of aging on development of disease pathology. Given that age is the greatest risk factor for AD, we expected to observe substantial and consistent acceleration of the tauopathy in the context of the aged mouse brain. While present, the degree of exacerbation did not seem very substantial, and was not present in all the markers we selected to examine. It is conceivable that had we chosen 3–4-month-old young mice to compare with mice of advanced age (20 month), we may have observed larger impacts of age on tauopathy. However, these extremes of age make it challenging to assess whether the differences result from maturation versus aging. Of some relevance is that although age increases risk of AD dementia, the pathology in older cases of AD is typically less severe than in younger ages [35 –37]. We conclude that the influence of age on tauopathy is present, but unlikely to account for the degree of increased risk for AD conferred by advanced age.
Footnotes
ACKNOWLEDGMENTS
We thank Nick Kanaan at Michigan State University for the donation of the TOC1 antibody and guidance of TOC1 ELISA methods, and Kevin Nash for the glycerol stock of GFP plasmid containing cells.
FUNDING
This work was supported by AG051500 to DM, AG062217 to MNG and the Spectrum Health-MSU Alliance Corporation.
CONFLICT OF INTEREST
None of the authors have a financial conflict of interest with the subject matter of this manuscript.
DATA AVAILABILITY
The data supporting the findings of this study are available on request from the corresponding author.