
Abstract
BACKGROUND:
It is now known regulatory effect of gaseous mediators in many bodily functions. These mediators include nitric oxide (NO), carbon monoxide (CO) and hydrogen sulfide (H2S). However, detailed data on the regulatory role of each of these gasotransmitters (GTs) are still not sufficiently studied.
OBJECTIVE:
The aim of this study was to investigate on models of microrheological behaviour of intact red blood cells and their recovered ghosts of cellular responses to all three known gasotransmitters: NO, H2S, and CO.
METHODS:
In experiments with intact red blood cells (RBCs) and their recovered ghosts (filled with an isotonic solution of known viscosity), deformability (RBCD) and aggregation (RBCA) were recorded after incubation of cells with GT donors or stimulators of their endogenous synthesis.
RESULTS:
It was found that all three GT donors moderately increased the deformability of both intact RBCs and their recovered ghosts (by 5–10%, p < 0.05). In addition GT donors and substrates of their endogenous synthesis significantly reduced RBCA, from 20 to 37% (p < 0.01). Experiments with inhibition of soluble guanylate cyclase (s-GC) almost completely eliminated the RBCD increasing effect of GT donors
CONCLUSION:
In present study it was demonstrated that all three known gaseous mediators: NO, CO and H2S cause moderate statistically significant positive alterations in RBC deformability as well as a noticeable decrease in RBC aggregation. This was confirmed both in experiments with gasotransmitter donors and stimulators of GT endogenous synthesis. In addition, the data obtained suggest that RBC microrheological responses to the actions of all three GTs are associated with activation of the guanylate cyclase signalling cascade.
Keywords
Introduction
Over the last few decades there has been intense interest in a group of gaseous mediators known as gasotransmitters (GT); currently comprising nitric oxide (NO), carbon monoxide (CO) and hydrogen sulfide (H2S) [1–3]. It is known that for efficient oxygen transport into tissues, red blood cells must have optimal microrheological characteristics [4–6]. It was shown that their cellular properties change positively under the influence of an NO donor, sodium nitroprusside, or upon activation of endothelial NO synthase (eNOS) by L-arginine [7, 8]. Although the biochemistry and physiology of NO have been widely studied, hydrogen sulfide (H2S) has only recently been recognized as an important signalling molecule [9, 10]. Hydrogen sulfide is an endogenous gaseous mediator affecting many physiological and pathophysiological conditions [3, 11].
Discoveries during the last decade on the endogenous synthesis of H2S in the mammalian system and its protective role in combating cellular necrosis, apoptosis, oxidative stress, inflammation as well as promoting angiogenesis and inhibits platelet aggregation and white blood cell adhesion [1, 12]. However, there is practically no data on the effect of hydrogen sulfide on the microrheology of the largest population of blood cells, on erythrocytes.
The low molecular weight gas carbon monoxide (CO), and similar gaseous molecules (e.g., H2S, nitric oxide) have been implicated as potential inhalation therapies in inflammatory diseases [13]. Carbon monoxide (CO) is an endogenously derived gas formed from the breakdown of heme by the enzyme heme oxygenase (HO). CO is now recognized as a key signalling molecule that regulates numerous cardiovascular functions [14, 15]. The release of CO by vascular cells exerts both paracrine and autocrine effects on vascular smooth muscle cells (SMC) and circulating blood cells and also reduces platelet aggregation [16, 17]. Accumulated experimental data indicate more and more clearly on the possible role of endogenous carbon monoxide (CO) in cell cytoprotection, and shows it as a probable factor participating in modulation of the vascular tonus in hypoxia [18]. It was shown that, for example, smooth muscle-derived CO may contribute to endothelium-independent regulation of vascular tone by providing a vasodilatory influence [19].
The aim of the study was to investigate the effect of three donors of gasotransmitters of nitric oxide, hydrogen sulfide, carbon monoxide and their substrates of endogenous synthesis on the red blood cell (RBC) microrheology.
Materials and methods
The study was approved by the local Ethics Committee of the University (protocol No. 6, 18.06.2020), and an informed consent of all the subjects were obtained according to the recommendations of the Declaration of Helsinki (The WMA’s Declaration of Helsinki on Ethical Principles for Medical Research Involving Human Subjects, as amended by the 64th WMA General Assembly, Fortaleza, Brazil, October, 2013).
Whole blood samples (9 mL) from healthy donors (n = 20) were drawn via venipuncture into vacuum tubes with EDTA (4.1 mM). The RBCs were separated from plasma by centrifugation (15 min at 3000 rpm), washed in isotonic sodium chloride solution supplemented with glucose (5.0 mM), and resuspended in the Ringer solution with the addition of dextran 200 (10% HAES-steril, Fresenius Kabi, Germany), to stimulate RBC aggregation. The viscosity of this fluid was 1.30 mPas and RBC suspension hematocrit (Hct) for microrheological measurement was 40.0±0.3%. To assess the effects of gasotransmitters (GTs) on the RBC deformability (RBCD) and their aggregation (RBCA) cells were incubated for 30 min at 37°C with one of the following substance: sodium nitroprusside (SNP, 100μM), as a donor of NO; L-arginine (100μM), substrate of endothelial NO synthase (eNOS); sodium hydrosulfide (NaHS, 100μM) as a hydrogen sulfide donor; L-cysteine with a substrate for the formation of hydrogen sulfide, cystionine-gamma-lyase (CSL, 100μM); Tricarbonylchloro(glycinato)ruthenium (II) (CORM–3, 50μM), as a donor of carbon monoxide (CO); Biliverdin hydrochloride (BV HCl, 1.0μM), as stimulator of hemoxygenase 1 (HO-1); 1H-[1, 4]- oxadiazole[4, 3-a]quinoxalin-l-one (ODQ, 1.0μM), a soluble guanylate cyclase (sGC) inhibitor; Only in Ringer’s solution (without drug) - control samples.
RBC suspension was incubated the same way (duration and temperature) but in a drug-free buffer solution was used as a control sample. The drugs were dissolved in DMSO or distilled water. To prepare recovered ghost of red blood cells 7 ml of distilled water cooled to 4° C were added to 1 ml of a washed red blood cell suspension (Hct = 95%) and then centrifuged for 25 minutes at 8,000 g. After removal of the supernatant to the bottom fraction of the centrifuge, 9 ml of an isotonic Ringer solution with dextran 200 were added and incubated 30 min at 37°C. The suspension of the recovered ghost of RBCs was divided into several aliquots and incubated with the aforementioned compounds. Their deformability was also recorded after the incubation.
Hemorheological measurements
Red blood cell aggregation was assessed by the Myrenne aggregometer which provides an index of RBC aggregation facilitated by low shear. In brief, the suspension was subjected to a short period of high shear to disrupt pre-existing aggregates, following which the shear was abruptly reduced to 3 s–1 and light transmission through the suspension that was integrated for ten seconds; the resulting index, termed “M1” by the manufacturer and “RBCA” herein, increased with enhanced RBC aggregation.
A parallel plate flow channel was used to estimate red blood cell deformability (for details see: [20]. In brief, the cells were attached to bottom part of the chamber with “one point” and then they were deformed by shear flow, under constant shear stress (2.0 N/m2). The length (L) and width (W) of each of about hundred cells were measured and red blood cell elongation index of individual red blood cells was calculated as an index of red blood cell deformability (RBCD) according to:
Miscellaneous techniques
Whole blood and red cell suspension hematocrits were determined via the microhematocrit methods (i.e., 12,000×g for 7 minutes).
Statistics and data presentation
The results are presented as mean±SEM. Comparisons were made using paired t-tests, and a two-tailed p value of p < 0.05 was considered to be significant; given the exploratory nature of these studies, p values are reported as determined.
Results
The effect of nitrogen oxide on red blood cell microrheology
Analysis of the data showed that the donor NO, SNP increased the deformability of red blood cells by 9% (from 1.94±0.02 to 2.11±0.01 units, p < 0.01). As for the substrate for e-NOS, L-arginine, after incubation with it, RBCD increased to 2.13±0.01 units, which is 10% (p < 0.01) more than without the drug (Fig. 1A).
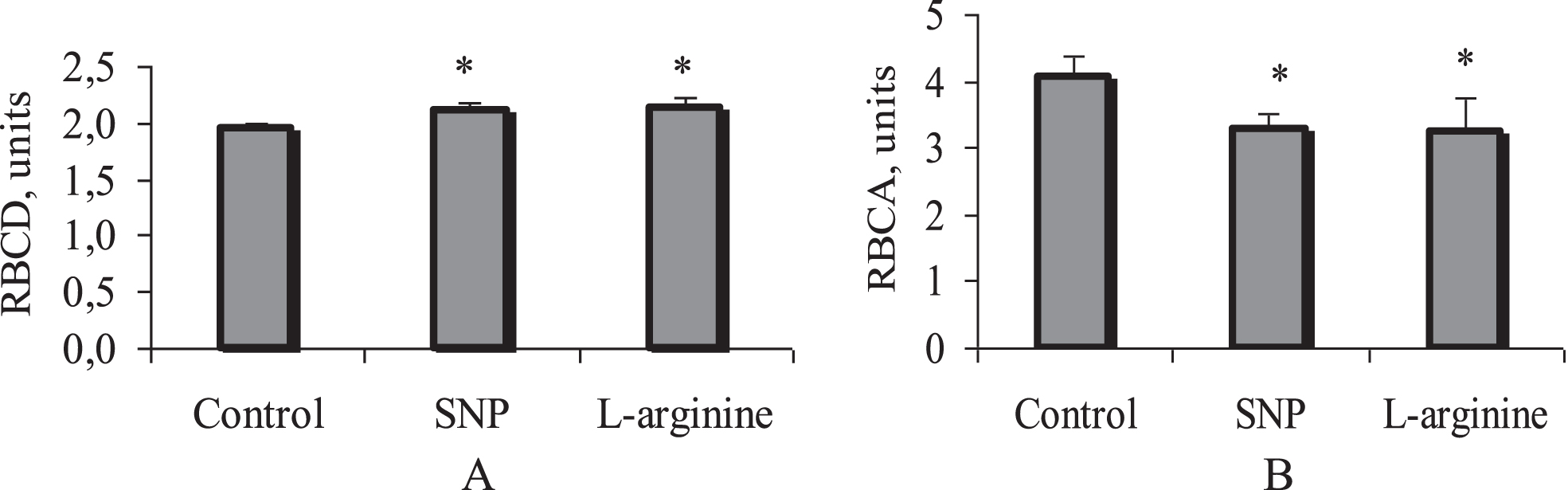
Alteration of red blood cell deformability (A) and aggregation (B) under the influence of incubation with sodium nitroprusside (SNP, 100μM) and L-arginine (100 μM). Note: RBCD –red blood cell deformability; RBCA –red blood cell aggregation; *p < 0.01, vs. control. n = 18 in each group.
The NO donor and the e-NOS substrate reduced RBCA approximately equally. After incubation with them, a decrease of RBCA was about 20–27% (p < 0.01) relative to the control level of this microrheological parameter (Fig. 2B). For example, cell incubation with SNP resulted in a RBCA decrease by 27% (from 4.16±0.11 to 3.02±0.08 units; p < 0.01).
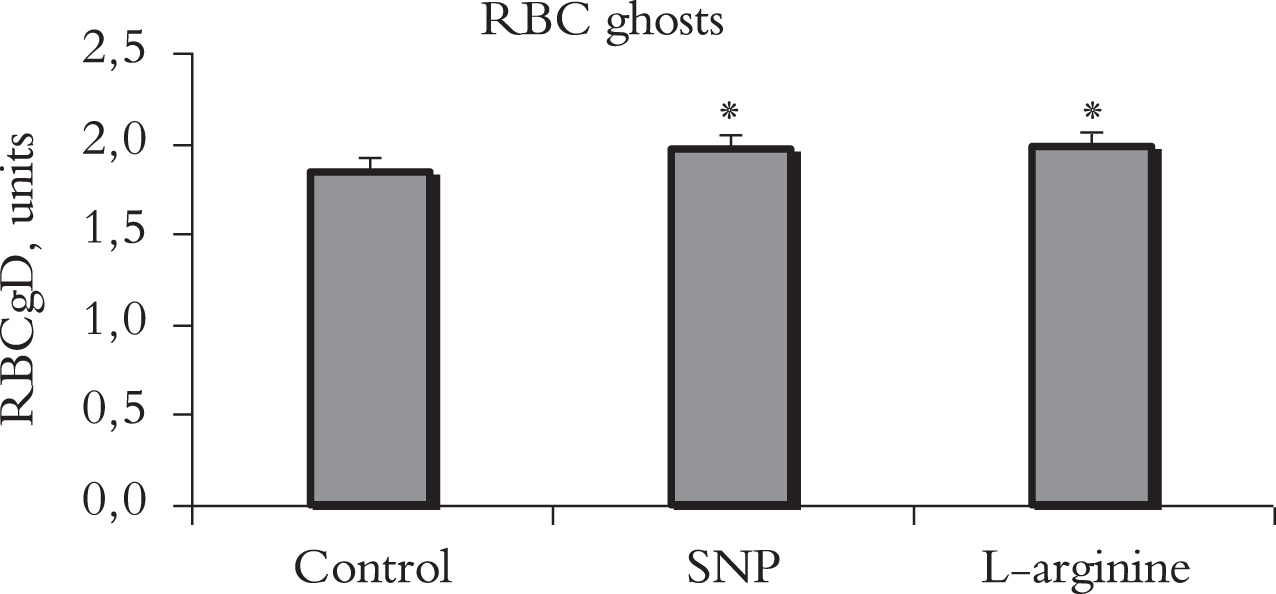
Alteration of red blood cell ghost deformability under the influence of incubation with sodium nitroprusside (SNP, 100μM) and e-NOS substrate, L- arginine (100μM). Note: RBCgD –red blood cell ghost deformability; *p < 0.01, vs. control. n = 18 in each group.
When analyzing the microrheological responses of the recovered RBC ghosts (RBCg) to the effect of the SNP a moderate increase in their deformability was found. The rise of this characteristic was 7% (from 1.84±0.03 to 1.96±0.02 units, p < 0.01).
When L-arginine, was added to the RBCg suspension, the increase of the ghost deformability did not differ much from the microrheological effect of SNP, an 8% increase was recorded (Fig. 2).
After RBC incubation with NaHS as a hydrogen sulfide donor RBCD was increased by 10% (from 1.97±0.04 to 2.16±0.03 units, p < 0.01). The microrheological effect of the CSE enzyme substrate L-cysteine was slightly less pronounced than after cell incubation with SNP (Fig. 3A). The RBCD increase was only 6% (from 1.97±0.04 to 2.09±0.02 units). However, this alteration was also statistically significant (Fig. 3A, p < 0.01).
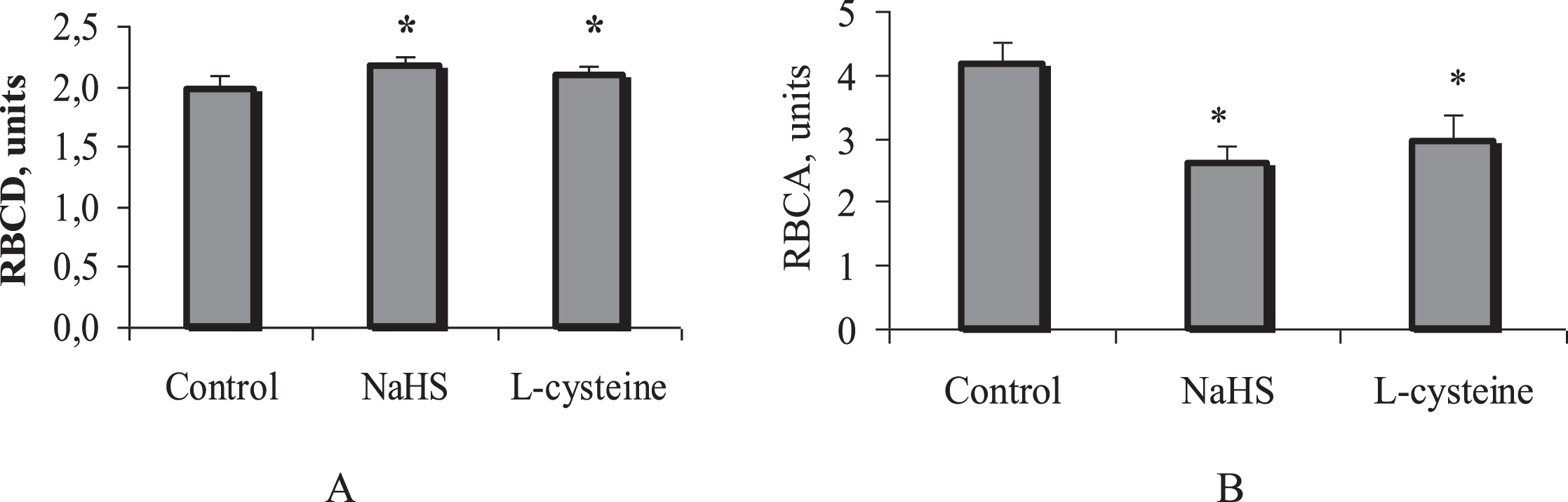
Alteration of red blood cell deformability (A) and aggregation (B) under the influence of incubation with sodium hydrosulfide (NaHS, 100μM) and L-cysteine (100μM). Note: RBCD –red blood cell deformability; RBCA –red blood cell aggregation; *p < 0.01, vs. control. n = 16 in each group.
A study of RBCA after cell incubation with NaHS showed that its decline was 37% (from 4.16±0.24 to 2.61±0.28 units, p < 0.01). Whereas the RBC aggregation-reducing effect of L-cysteine was 29% (from 3.81±0.23 to 2.71±0.32 units, p < 0.01; Fig. 3B).
The deformability of RBC ghosts was increased by 9% (Fig. 4) after their incubation with a hydrogen sulfide donor (from 1.83±0.02 to 1.99±0.03 units, p < 0.01).
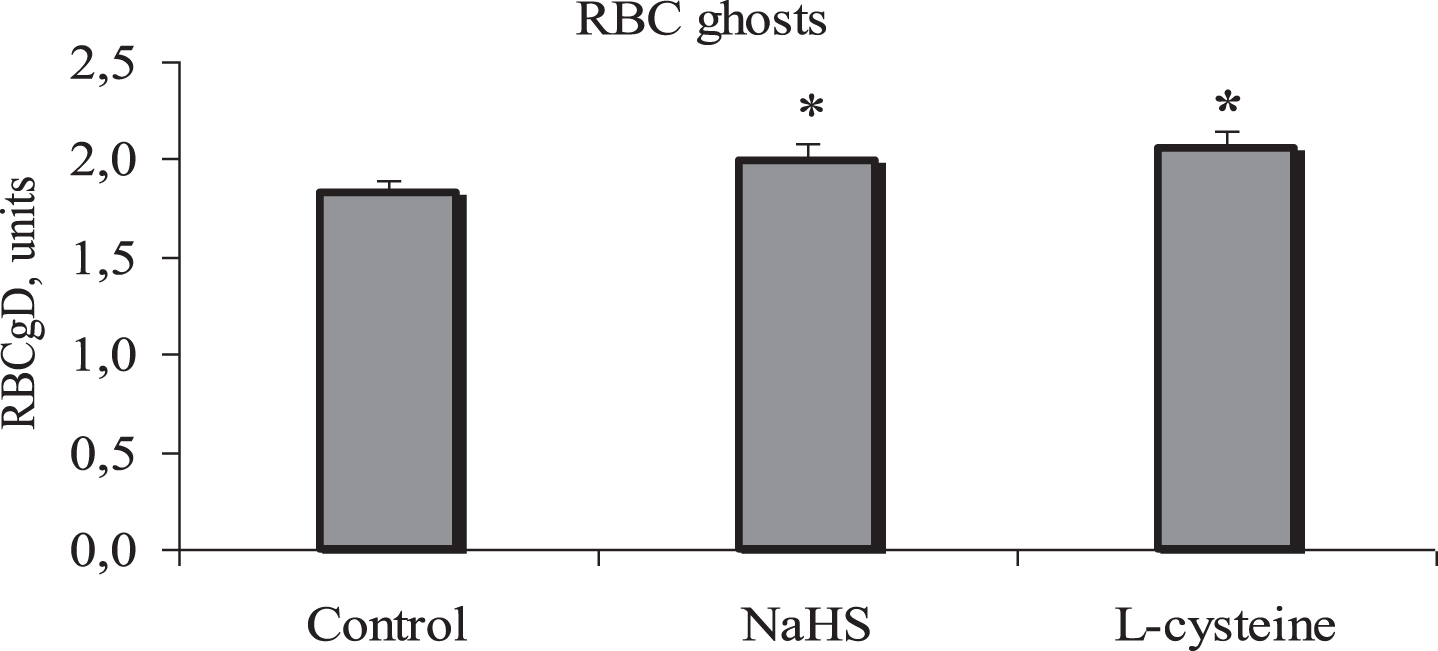
Alteration of red blood cell ghost deformability under the influence of incubation with a hydrogen sulfide donor (NaHS, 100μM) and cystathionine gamma-lyase (CSL) stimulator, L- cysteine (100μM). Note: RBCgD –red blood cell ghost deformability; *p < 0.01, vs. control. n = 16 in each group.
L-cysteine more significantly changed the deformability of RBCg. In this case, the deformability increase reached 12% (from 1.83±0.02 to 2.05±0.03 units) and was also statistically significant (p < 0.01, Fig. 4).
RBC incubation with CORM-3 as a CO donor was accompanied by a significant increase in RBC deformability (Fig. 5A). Their elongation index increased from 2.01±0.01 to 2.12±0.01 units (p < 0.01).
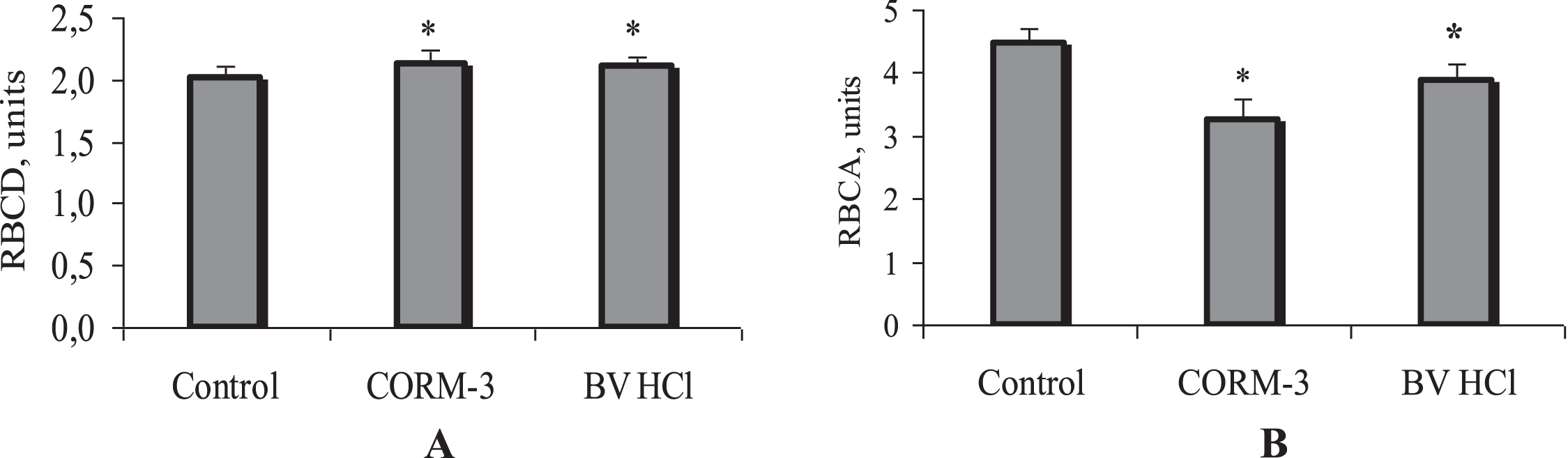
Alteration of red blood cell deformability (
The heme oxygenase stimulator, biliverdin, also contributed to an increase in RBCD up to 2.10±0.02 units, p < 0.01). This increase in RBC deformability by 5% (p < 0.01) was slightly less than after incubation of cells with CORM-3, but it was also statistically significant (Fig. 5A).
As regards the aggregation of red blood cells, the CO donor significantly reduced it by 27% (from 4.46±0.26 units –control to 3.26±0.18 units –CORM-3). Incubation of cells with biliverdin was accompanied by a moderate decrease in RBCA, by 13% (from 4.37±0.14 to 3.80±0.12 units, p < 0.05; Fig. 5B).
After RBCg incubation with a CO donor, CORM-3 a 10% increase in deformability was observed (from 1.87±0.01 to 2.05±0.02 units, p < 0.01).
If the red blood cell ghosts were incubated with biliverdin chloride, then their deformability increased by 9% (p < 0.01; Fig. 6): from 1.87±0.01 to 2.04±0.02 units.
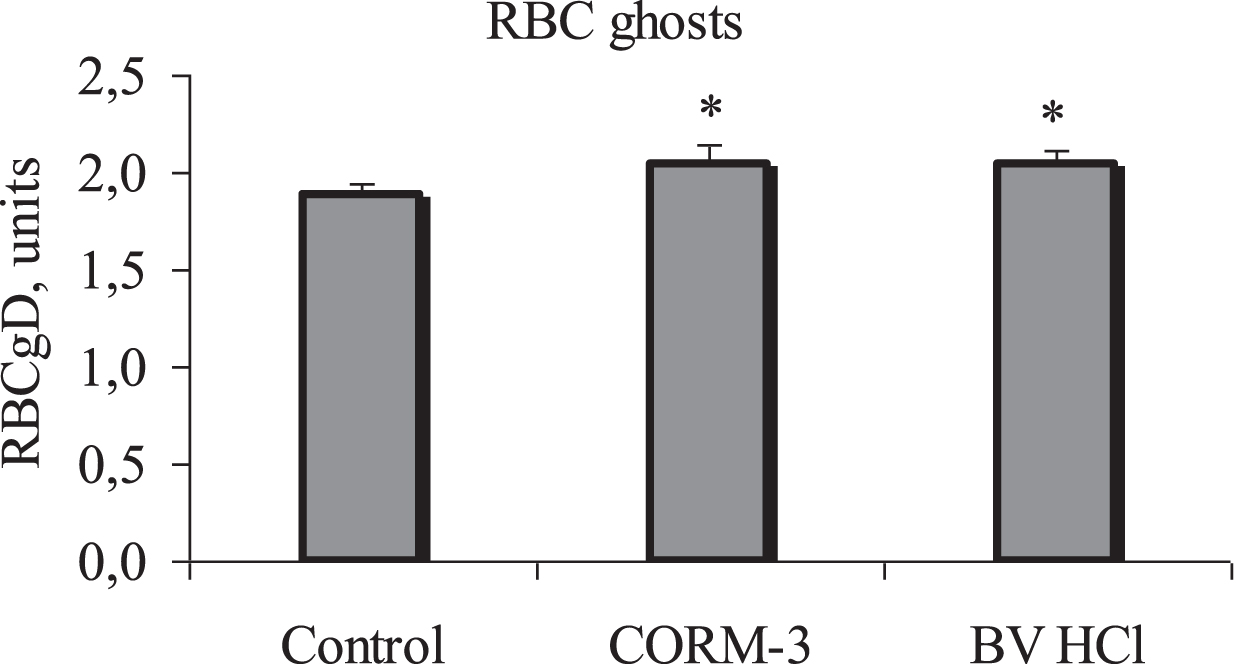
Alteration of red blood cell ghost deformability under the influence of incubation with a carbon monoxide donor (CORM-3, 50μM) and heme oxygenase activity stimulator, biliverdin hydrochloride (50μM). Note: RBCgD –red blood cell ghost deformability; BV HCl –Biliverdin hydrochloride; *p < 0.01, vs. control. n = 12 in each group.
The results of the study showed that both the action of the nitric oxide donor and the stimulation of its endogenous synthesis in red blood cells led to a positive change in the red blood cell microrheological properties. The effect of the two compounds was very similar. Thus, RBC deformability increased by 9-10%, and their aggregation decreased by 20% in both series of experiments. These observations can confirm the opinion about the possibility of endogenous synthesis of NO in mature red blood cells [4, 20]. It is believed that the main molecular target for NO in cells is soluble guanylate cyclase (s-GC) [22, 23]. However, in experiments with red blood cell ghosts (RBCg), where the cytoplasm and, consequently, s-GC, were removed, a significant increase in their deformability in response to a NO donor and a stimulator of e-NOS activity was shown. In both series of experiments, an increase in RBCg deformability by 7-8% was observed (p < 0.01). These microrheological responses of whole red blood cells and their recovered ghosts (where there is no s-GC) to nitric oxide may indicate that there are probably other intracellular signalling pathways for the implementation of these responses. It is known that this GT can activate the c-GMP-independent regulatory mechanism [24]. For example, it has been shown that NO modifies ATP-sensitive K+ channels [25], and this is accompanied by positive vascular changes and RBC microrheology [26, 27]. Furthermore nitrosylation and transnitrosylation of RBC membrane and cytoskeletal proteins are also possible [28].
S-nitrosation of intracellular proteins such as haemoglobin or especially spectrin can alter membrane viscoelasticity and the deformability of the whole cell [29]. Therefore, it can be assumed that the positive microrheological effects of NO in RBCs are realized by more than one molecular signalling pathway.
NaHS, the donor of another gasotransmitter, hydrogen sulfide, influenced the deformability of intact RBCs and their recovered ghosts like SNP or L-arginine. At the same time its effect on RBCD was slightly greater than that of L-cysteine. As for the RBC ghost, their deformability changed a little more noticeable under the influence of L-cysteine than NaHS. It is known that ATP-dependent potassium channels are considered as a cell molecular target of hydrogen sulfide [25, 30]. However, the blocking of these channels with glibenclamide did not remove the increase in RBC deformability under the influence of sodium hydrosulfide [27]. Therefore, it can be assumed that hydrogen sulfide, as a gaseous mediator, uses a different signalling pathway.
The obtained data indicate that the carbon monoxide donor CORM-3 and the heme oxygenase (HO) activator, biliverdin hydrochloride (BV HCl) less markedly increased RBC deformability than donors NO and H2S. Like two other GT donors, CORM-3 reduced RBC aggregation by a similar value of 27% (p < 0.01). CO-releasing molecule-3 (CORM-3) resulted in increased RBC ghost deformability by 10% (p < 0.01). A similar change in the RBCg deformability caused BV HCL. It can be assumed that these compounds activate the metabolism of carbon monoxide and have a moderate effect on the red cell microrheology. It is known that constitutive heme oxygenase cleaves heme to form biliverdin and carbon monoxide (CO) [13, 31]. In turn, biliverdin activates HO and promotes the formation of CO in cells [32].
In is known that the NO- and CO-induced a change in cellular functions are mainly mediated by the cGMP pathway [33]. Therefore CO can stimulate s-GC to increase the production of guanosine 3’5’-monophosphate (cGMP), which has been demonstrated in vascular smooth muscle cells [34]. However, there is an opinion that another GT, hydrogen sulphide can realize its effects independent of the activation of the cGMP pathway [35]. The authors showed that the CO-induced vasorelaxation was not inhibited by ODQ. On a model of RBC microrheological response, we found that upon preliminary incubation of RBCs with ODQ as an s-GC inhibitor, a similar RBC deformability enhancing effect of all three gasotransmitter donors was almost completely eliminated (Fig. 7).
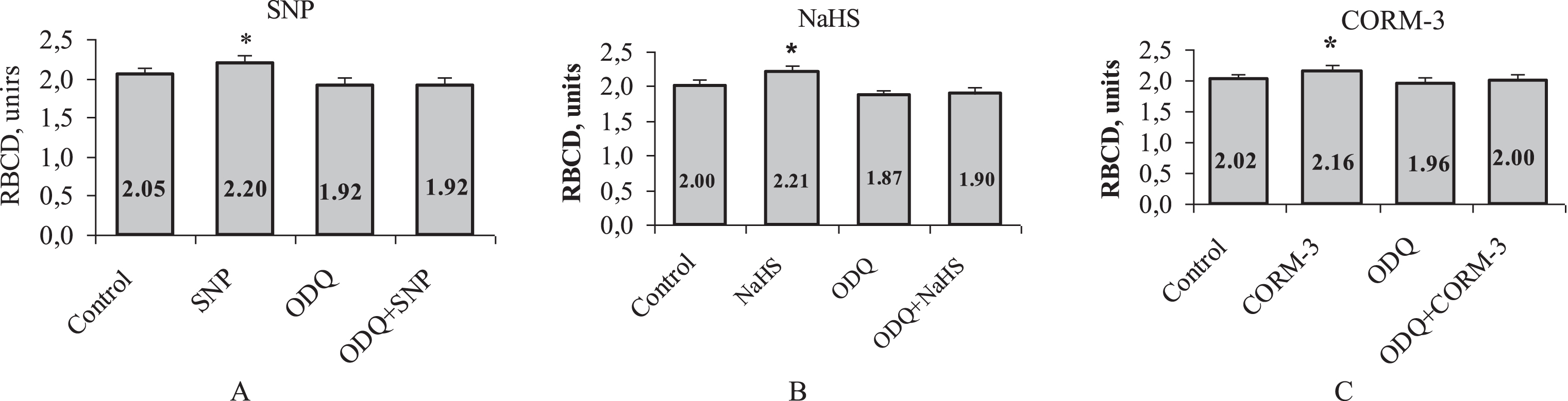
Alterations of red blood cell deformability under the influence of three gasotransmitter donors (A, B, C) after soluble guanylate cyclase (s-GC) inhibition with 1H-[1,2,4, 1,2,4]- oxadiazole [4, 3-a]quinoxalin-l-one (ODQ, 0.5μM). *p < 0.01 relative to control. n = 12 in each group. Note: SNP –sodium nitroprusside; NaHS –sodium hydrosulfide; CORM-3 –tricarbonylchloro(glycinato)ruthenium (II).
Our novel observations showed that the inhibition of cGMP signalling pathway eliminates the influence of donors of all three gasotransmitters on the microrheological responses of intact red blood cells. The use of ODQ as the specific s-GC inhibitor to solve this problem suggests that this particular enzyme can be a molecular target for the action of all three gasotransmitters.
In conclusion, in present study, it was demonstrated that all three known gaseous mediators: nitric oxide, carbon monoxide and hydrogen sulfide cause moderate statistically significant changes in the red blood cell deformability and their recovered ghosts, as well as a noticeable decrease in red blood cell aggregation. This was confirmed both in experiments with donors of gasotransmitters and with stimulants of enzymes of their endogenous synthesis. The similarity in the direction of the changes and the magnitude of the microrheological responses of RBCs to GT donors suggests that all three gasotransmitters can use the same molecular signalling pathway. Experiments with inhibition of soluble guanylate cyclase eliminated the effects of donors of all three GTs increasing the deformability of red blood cells. Thus, we can assume that all three gasotransmitters use the same signalling pathway associated with the activation of guanylate cyclase-cGMP signalling cascade.
Footnotes
Acknowledgments
The reported study was funded by RFBR according to the research project No 18-015-00475-a.
Conflict of interest
The authors have no conflict of interest to report.