
Abstract
Exposure to noise trauma, such as that from improvised explosive devices, can lead to sensorineural hearing loss and a reduced quality of life. In order to elucidate the mechanisms underlying noise-induced hearing loss, we have adapted optical coherence tomography (OCT) for real-time cochlear visualization in live mice after blast exposure. We demonstrated that endolymphatic hydrops develops following blast injury, and that this phenomenon may be associated with glutamate excitotoxicity and cochlear synaptopathy. Additionally, osmotic stabilization of endolymphatic hydrops partially rescues cochlear synapses after blast trauma. OCT is thus a valuable research tool for investigating the mechanisms underlying acoustic trauma and dynamic changes in endolymph volume. It may also help with the diagnosis and treatment of human hearing loss and/or vertigo in the near future.
Keywords
Introduction
Exposure to noise trauma, such as that from improvised explosive devices, can cause significant hearing impairment among military personnel and civilians [3, 10]. A 2007 study showed that 58 percent of military personnel who suffer blast-related injuries also demonstrate hearing loss, resulting in a significant reduction in quality of life [2, 29]. It is therefore important to understand the mechanisms underlying acoustic trauma in order to develop therapies for this common cause of disability.
While hair cell damage is the most common explanation for hearing loss following traumatic noise exposure, another proposed mechanism is a loss of cochlear synapses. Synaptopathy results in decreased communication between hair cells and auditory neurons, which may contribute to perceptual hearing abnormalities including tinnitus, hyperacusis and difficulty discerning speech-in-noise [16, 20]. Since these changes can only be measured in vitro, a direct means of assessing cochlear trauma in vivo is still necessary to further clarify the mechanisms underlying noise-induced hearing loss.
We have thus adapted optical coherence tomography (OCT) for real-time visualization of the cochlea in live mice. Using OCT, we have demonstrated that endolymphatic hydrops develops following blast exposure and that this phenomenon is associated with cochlear synaptopathy [12]. Additionally, we have found that endolymph volume can be modulated through round-window application of solutions with varying tonicities and that this technique partially rescues cochlear synapses after blast exposure. This manuscript reviews and summarizes our findings from previously published work [12]. Here, we expand upon our previously reported findings and describe how OCT, our study methodology, and the interpretation of our results elucidate one possible mechanism for the generation of endolymphatic hydrops and its effect on the cochlear and vestibular systems [12]. We believe that OCT is a valuable research tool for investigating these systems in animal models and may be a useful imaging modality for understanding dynamic changes in endolymph volume in humans in the near future.
Review
Optical coherence tomography permits cochlear visualization in live mice
OCT is a noninvasive three-dimensional imaging technique that can visualize biological microstructures in vivo with a spatial resolution at the level of several micrometers [33]. It provides real-time subsurface imaging by directing a light beam onto tissue and measuring backscattered light as a function of depth [8]. The backscattered light is used to create cross-sectional images, with small changes in refractive index providing contrast [9]. Through segmentation of multiple OCT cross-sections, three-dimensional reconstructions of biological tissues can be rendered and analyzed.
OCT has been utilized for noninvasive diagnostic imaging in many tissues, including the eyes, coronary arteries, and skin [23]. Recently, there has been growing interest in using OCT to visualize the inner ear microanatomy. Many conditions causing hearing loss and vertigo cannot be effectively studied because the cochlea is too small to be visualized by traditional imaging techniques. This makes OCT a valuable tool for diagnosing inner ear disorders. Figure 1A-B demonstrate the cochlear substructures which can be visualized using OCT.
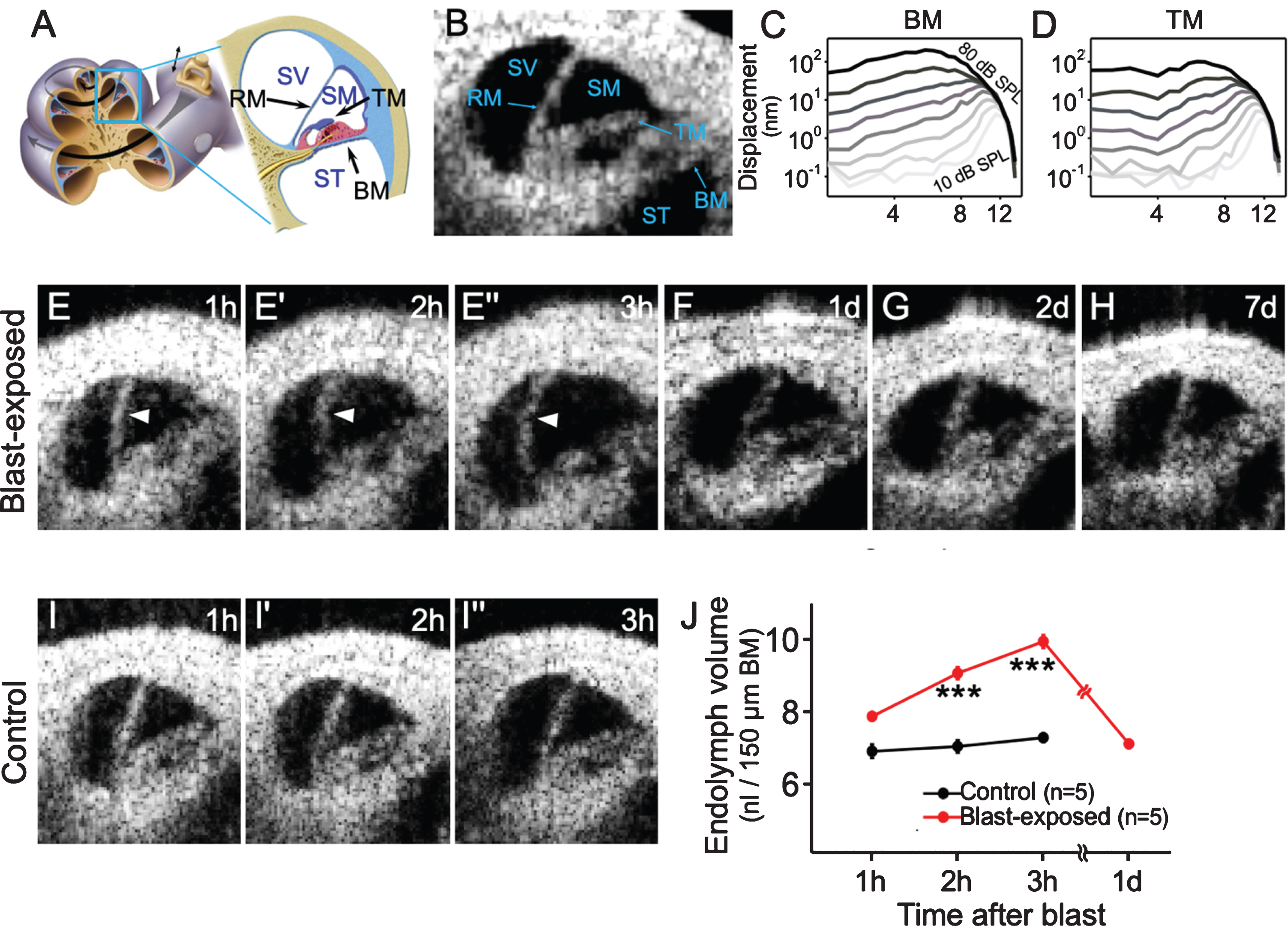
Blast exposure causes endolymphatic hydrops. (A) Illustration of the mouse cochlea highlighting the apical turn where optical coherence tomography (OCT) images are obtained. (B) Cross-sectional OCT image of the cochlea in a control mouse with labeled substructures. (C, D) Normal nonlinear vibratory responses measured from the basilar membrane (BM) and tectorial membrane (TM) of a representative control mouse display tuned responses at ∼12 kHz. (E) Endolymphatic hydrops (arrowheads) developed within the first 3 h following blast exposure. (F– H) OCT images 1, 2, and 7 d after blast exposure showed normal endolymph volume with no change in the position of Reissner’s membrane (RM). (I) OCT images from a representative live control mouse showed normal endolymph volume over 3 h. (J) Endolymph volume in blast-exposed mice was increased compared with unexposed control mice. SV, scala vestibuli; SM, scala media; ST, scala tympani. ***P < 0.001. Figure adapted from [12].
We have adapted OCT to not only visualize the mouse cochlea, but also to measure vibrations of cochlear microstructures using a technique called Volumetric Optical Coherence Tomography and Vibrometry (VOCTV) [9, 17]. This system assesses cochlear function by measuring sound-induced subnanometer vibrations throughout the organ of Corti. Vibrational measurements of different microstructures in a normal cochlea, including the basilar membrane (BM) and tectorial membrane (TM), show sharp tuning and large gains (Fig. 1C-D). Using VOCTV, we are able to detect changes in vibratory responses, such as the loss of cochlear amplification, indicating impaired cochlear function.
In order to elucidate the mechanisms underlying blast-induced hearing loss, we designed a blast chamber to create pressure waves similar to an improvised explosive device [3]. In CBA/CaJ mice exposed to a blast wave with the strength of a roadside bomb (peak pressures 130±9 kPa [∼196 dB SPL]), sequential OCT imaging showed progressive bulging of Reissner’s membrane in the first three hours following blast exposure (Fig. 1E). This suggested an increase in endolymph volume, or endolymphatic hydrops [12]. By 24 hours following blast exposure, Reissner’s membrane returned to its original position where it remained for at least one week (Fig. 1F– H). Quantification of endolymph volume within the scala media demonstrated that blast exposure resulted in increased endolymph volume which returned to baseline by 24 hours after the blast (Fig. 1J).
Changes in K+ cycling underly the development of blast-induced endolymphatic hydrops
Alterations in K+ cycling may play a role in the development of endolymphatic hydrops after blast exposure. Ordinarily, K+ is continuously secreted into the endolymph by the stria vascularis and removed through stereociliary transduction channels and other pathways [35, 38]. It is therefore possible that blast-induced damage to outer hair cell (OHC) stereocilia impairs K+ resorption through apical transduction channels. Water is then driven into the endolymph by the increased osmotic load due to the elevated concentration of K+ [12].
To test this hypothesis, we utilized scanning electron microscopy to evaluate the stereocilia of CBA/CaJ mice sacrificed immediately after blast exposure. In contrast to unexposed mice, blast-exposed mice demonstrated hair bundle damage throughout the cochlear base with stereocilia that were fused together, bent or missing (Fig. 2A-B) [12]. Thus, blast exposure appeared to cause immediate stereociliary damage.
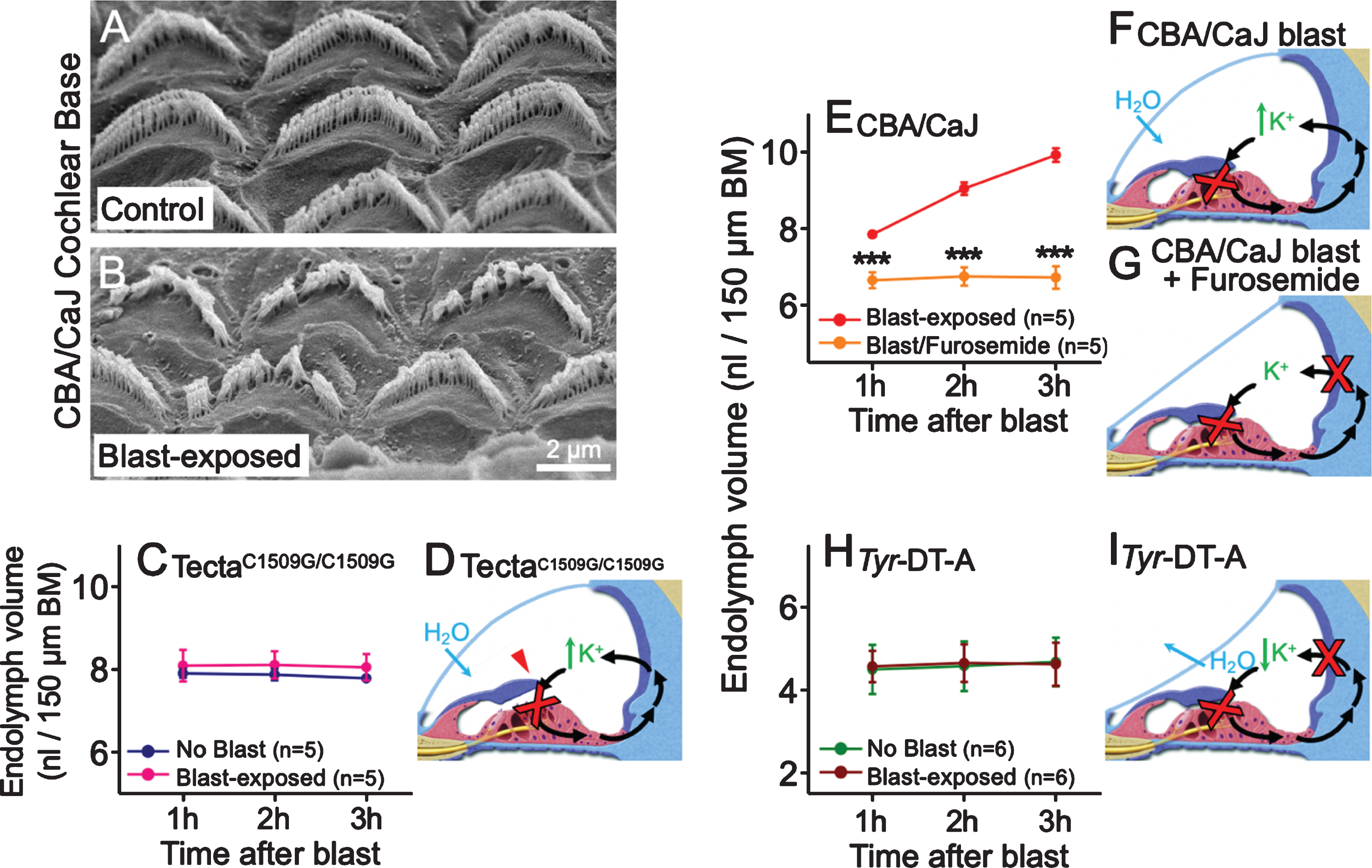
Stereociliary trauma and dysregulation of K+ cycling are necessary for the development of endolymphatic hydrops after blast exposure. (A, B) Scanning electron microscopic images show damage to OHC stereociliary bundles in the cochlear base of blast-exposed CBA/CaJ mice compared with no damage in unexposed control mice. (C) Endolymph volume was higher than normal but unaffected by blast exposure in TectaC1509G/C1509G mice. (D) The TM is elevated off the epithelium in TectaC1509G/C1509G mice, so stereocilia are not sheared by the TM after blast exposure. K+ bias currents through transduction channels (red X) are reduced at baseline due to lack of static displacement of the stereociliary bundles, leading to elevated endolymph volume. However, K+ bias currents are not further reduced after blast exposure, so endolymphatic [K+] and fluid volume are expected to remain stable. (E) Endolymphatic hydrops develops in CBA/CaJ mice exposed to blast but not in mice pretreated with furosemide. (F) In CBA/CaJ mice exposed to blast, damaged stereocilia reduce K+ bias currents through transduction channels (red X), increasing endolymph [K+] and drawing in H2O. This leads to endolymphatic hydrops. (G) After furosemide treatment, K+ secretion by the stria vascularis (red X) is also blocked, so endolymphatic [K+] and fluid volume are expected to remain stable. (H) Endolymph volume in Tyr-DT-A mice was lower than normal, yet unchanged, by blast exposure. (I) Damaged stereocilia from blast exposure reduce K+ bias currents through transduction channels (red X) in Tyr-DT-A mice, but K+ secretion is also blocked due to ablation of stria intermediate cells (red X) in these mice. Thus, endolymphatic [K+] and volume are expected to remain stable. BM, basilar membrane. ***P < 0.001. Figure adapted from [12].
We then studied TectaC1509G/C1509G mutant mice, in which the TM does not deflect OHC stereocilia because it is detached from the organ of Corti [37]. Endolymph volume in these mice did not increase after blast exposure (Fig. 2C-D) [12]. The findings suggested that blast-induced stereociliary trauma occurs when stereocilia are sheared by the TM, and that this trauma is necessary to produce endolymphatic hydrops.
Next, CBA/CaJ mice were injected with furosemide before blast exposure to inhibit the Na-K-Cl cotransporter which secretes K+ into the scala media [1]. These mice did not develop blast-induced endolymphatic hydrops (Fig. 2E-G). We also investigated Tyr-DT-A mice, in which ablation of stria intermediate cells abolishes K+ secretion from the stria vascularis [11]. Similar to mice pretreated with furosemide, endolymphatic hydrops was not observed (Fig. 2H-I). Taken together, these results are consistent with the idea that blast-induced endolymphatic hydrops occurs because the rate of K+ resorption through damaged stereociliary trans-duction channels is reduced relative to its secretion by the stria vascularis [12].
Beyond changes in K+ cycling, alterations in perilymph tonicity also affect endolymph volume. Round-window delivery of therapeutics to the cochlea has been established as an effective clinical technique [4, 36]. Thus, we attempted to manipulate perilymph tonicity through middle ear injection of solutions with varying osmolarities (Fig. 3A). Round-window application of a solution that was hypotonic (distilled water, 0 mOsm/kg) relative to the perilymph and endolymph forced water into the scala media, causing Reissner’s membrane to bulge outwards and endolymphatic hydrops to develop. By contrast, round-window application of a hypertonic solution (artificial perilymph, 1,532 mOsm/kg) had the opposite effect, causing Reissner’s membrane to shift inwards and endolymph volume to decrease. Application of a normotonic solution (artificial perilymph, 304 mOsm/kg) had no effect on endolymph volume [12].
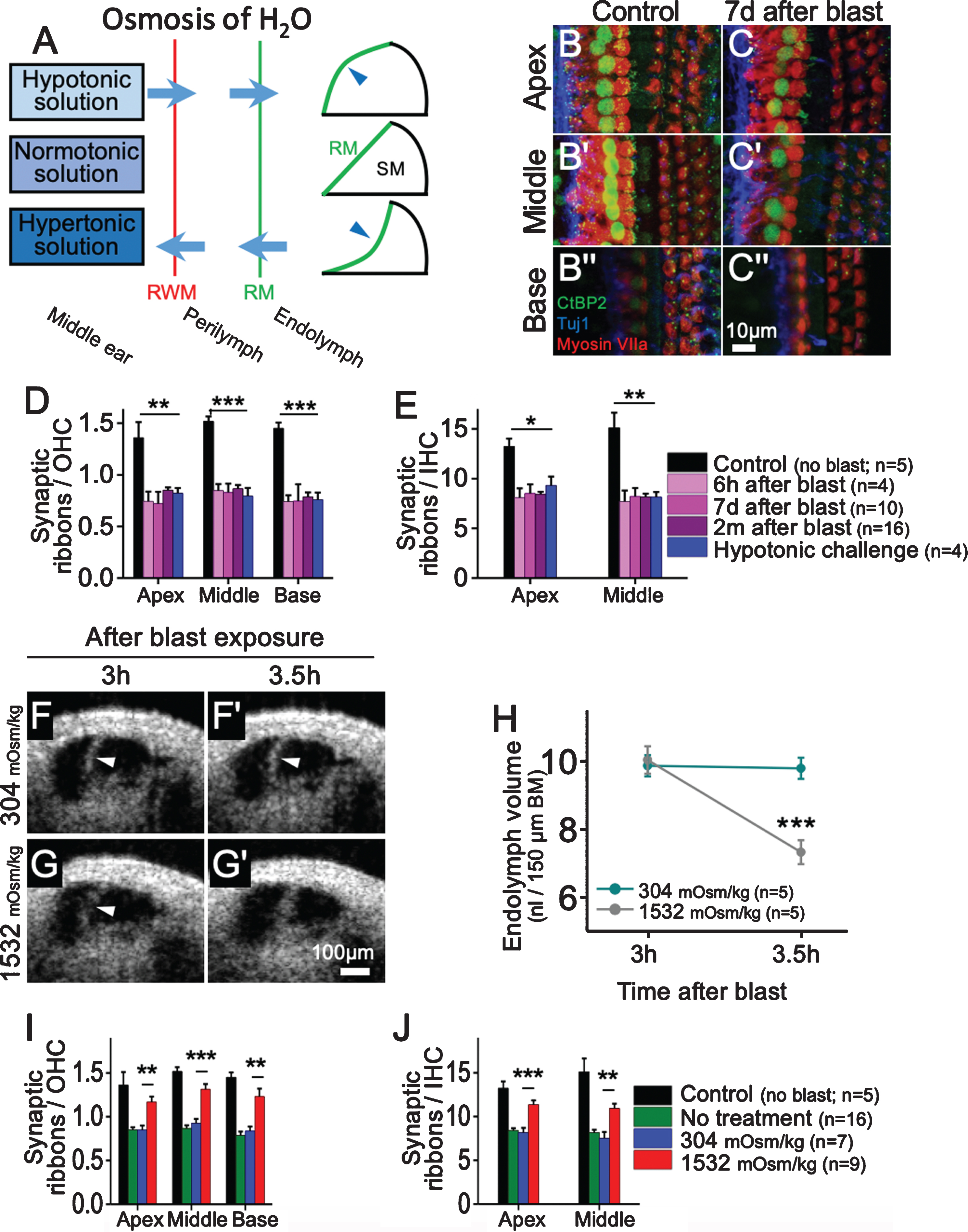
Blast exposure and hypotonic challenge cause cochlear synaptopathy, while hypertonic treatment partially prevents blast-induced synaptopathy. (A) Simple model of osmotic diffusion through the round window membrane (RWM) to alter endolymph volume and shift Reissner’s membrane (RM). A hypotonic solution applied to the round window membrane causes endolymphatic hydrops, while a hypertonic solution reduces endolymph volume. A normotonic solution has no effect. (B, C) The organ of Corti 7 d after blast exposure. Immunolabeling was utilized to visualize synaptic ribbons (CtBP2), auditory neurons (Tuj1), and hair cells (myosin VIIa). (D, E) Quantification of synaptic ribbons per OHC and IHC in control mice, blast-exposed mice, and mice treated with hypotonic challenge. Compared with control mice, blast-exposed mice and those receiving round-window application of a hypotonic solution had similar reductions in synaptic ribbons per OHC and IHC throughout the cochlea. (F, G) Representative mice were treated with round-window application of normotonic (304 mOsm/kg) or hypertonic (1,532 mOsm/kg) artificial perilymph 3 h after blast exposure. Repeat images were taken 30 minutes later. Mice treated with hypertonic artificial perilymph demonstrated an inward shift of Reissner’s membrane back to baseline, while those treated with normotonic perilymph did not. (H) Hypertonic artificial perilymph normalized endolymph volume, while normotonic treatment had no effect. (I, J) Quantification of synaptic ribbons per OHC and IHC in control mice as well as blast-exposed mice receiving no treatment, normotonic treatment, or hypertonic treatment. Hypertonic treatment partially rescued synaptic ribbon loss throughout the cochlea. BM, basilar membrane. *P < 0.05, **P < 0.01, ***P < 0.001. Figure adapted from [12].
To further characterize the cochlear damage caused by blast injury, we assessed inner hair cells (IHCs), OHCs, and ribbon synapses within the organ of Corti using standard immunolabeling protocols [12, 19]. One week after blast exposure, OHC death occurred throughout the cochlear base while IHCs were unaffected. To evaluate for cochlear synaptopathy, we counted the number of synaptic ribbons per OHC and IHC at multiple time points after blast exposure [16]. Throughout the cochlea, synaptic ribbons per OHC and IHC were significantly reduced in blast-exposed mice when compared with unexposed animals (Fig. 3B-E) [7, 12]. The pattern of basal OHC loss following noise trauma likely results from the large forces applied to the stereocilia of basal OHC and their increased metabolic vulnerability [5, 22]. Since there is no obvious mechanism explaining why the pattern of widespread synaptopathy does not match the more localized OHC death following blast exposure, it is possible that endolymphatic hydrops causes damage to synapses [12].
Endolymphatic hydrops is associated with glutamate excitotoxicity and synaptopathy
In order to determine if endolymphatic hydrops in the absence of blast injury leads to a loss of ribbon synapses, we applied a hypotonic solution to the round window of mice to induce endolymphatic hydrops. One week later, we counted hair cells and synaptic ribbons. Synaptic ribbons per IHC and OHC were reduced throughout the cochlea following the hypotonic challenge to a similar degree as after blast (Fig. 3D-E). However, OHC death was not observed following the hypotonic challenge [12]. Thus, isolated endolymphatic hydrops leads to cochlear synaptopathy without OHC death.
Prior studies have shown that glutamate excitotoxicity is responsible for the loss of synapses following noise trauma [26, 31]. To determine if glutamate excitotoxicity was the mechanism linking endolymphatic hydrops with cochlear synaptopathy, we utilized the drug CNQX, which blocks postsynaptic AMPA-type glutamate receptors [18]. Synaptic ribbons per IHC and OHC did not decrease when postsynaptic glutamate receptors were blocked with CNQX prior to the hypotonic challenge [12]. These results demonstrated that endolymphatic hydrops leads to cochlear synaptopathy through glutamate excitotoxicity.
We also evaluated whether isolated endolymphatic hydrops affects the cochlear amplifier. After inducing endolymphatic hydrops through round-window application of a hypotonic solution, we used VOCTV to measure the vibratory responses of the BM and TM. Vibratory response curves and gains were similar to unexposed mice, indicating that isolated endolymphatic hydrops does not alter OHC-mediated amplification [12].
Osmotic stabilization of endolymphatic hydrops partially rescues cochlear synapses
Since round-window application of a hypertonic solution reduced endolymph volume in control mice, we sought to determine if it could reverse endolymphatic hydrops in blast-exposed mice. Therefore, we applied hypertonic artificial perilymph to the round window of CBA/CaJ mice three hours after blast exposure once endolymphatic hydrops had developed. Endolymph volume returned to baseline within 30 minutes after hypertonic treatment (Fig. 3F– H) [12].
We then utilized immunolabeling to determine if hypertonic treatment could prevent blast-induced synaptopathy. Immediately after blast exposure, hypertonic or normotonic artificial perilymph was injected into the middle ear of CBA/CaJ mice, and two months later, hair cells and synaptic ribbons were counted. Basal OHCs were reduced to a similar degree in untreated mice and those undergoing normotonic or hypertonic treatment. However, mice treated with hypertonic artificial perilymph had significantly more synaptic ribbons per IHC and OHC following blast exposure when compared with untreated mice and those receiving normotonic treatment (Fig. 3I, J) [12]. Therefore, osmotic stabilization of endolymph with hypertonic treatment partially rescues synaptic ribbon loss following blast exposure.
Discussion
We have demonstrated that endolymphatic hydrops may be play a role in the development of cochlear synaptopathy following blast exposure. Trauma to OHC stereocilia is the first step of the process. Endolymphatic hydrops then develops, likely because K+ resorption from the endolymph is impaired due to stereociliary damage. In addition, hair cells are overstimulated and release excess glutamate. The resulting excitotoxicity damages synapses between hair cells and auditory nerve dendrites, leading to a loss of synaptic ribbons within hair cells. It is still unclear whether stereociliary damage occurs throughout the cochlea or just in the basal regions. Moreover, the question remains of how hair cells can be overstimulated if their stereocilia are damaged to the point that K+ is accumulating in scala media. Therefore, we postulate that tremendous K+ and Ca+2 influx into hair cells occurs during the blast exposure, and that this starts the process of glutamate excitotoxicity prior to the accumulation of K+ in the scala media.
Osmotic stabilization of endolymphatic hydrops through round window application of hypertonic treatment can partially ameliorate blast-induced cochlear synaptopathy [12]. This middle ear injection technique is commonly used in the clinic and could potentially be used on the battlefield to prevent cochlear damage in soldiers [6]. Intratympanic administration of a hypertonic solution may also have potential for treating Meniere’s disease, a disorder in which endolymphatic hydrops causes episodic vertigo, tinnitus, and sensorineural hearing loss [25].
Since OCT can resolve the morphology and vibratory responses of cochlear structures, it represents a potentially useful diagnostic tool in humans [14]. We have previously reported the use of OCT for in vivo middle ear imaging through the human tympanic membrane [13, 15]. OCT may also be useful for detecting endolymphatic hydrops in patients, which could help with predicting cochlear synaptopathy after noise trauma as well as diagnosing Meniere’s disease [24]. Additionally, OCT may prove valuable for guiding cochleostomies and electrode insertion during cochlear implantation [32].
Accessing the cochlea is the main challenge with adapting OCT for human use. While the mouse cochlea can be directly imaged through the otic capsule, the thickness of the human otic capsule prevents a similar approach. To date, OCT imaging of the human cochlea has only been reported in vitro [21, 32]. Therefore, we are working to develop an endoscopic OCT system to image the cochlea through the round window membrane in patients [14]. Hopefully, such a system will enable clinicians to make functional and morphological assessments of the inner ear, improving their ability to diagnose a variety of hearing and balance disorders.
Footnotes
Acknowledgments
Supported by R01 DC017741, R01 DC014450, and R01 EB027113.