
Abstract
Campylobacter spp. is a leading cause of community-acquired gastroenteritis, with rising antimicrobial resistance (AMR) posing a significant public health challenge. This study aimed to characterize the genomic basis of resistance and virulence in Campylobacter spp. isolated from human stool at the University Hospital of Split, Croatia. A total of 115 unduplicated isolates (100 Campylobacter jejuni and 15 Campylobacter coli) were identified and tested for antimicrobial susceptibility, from January to June 2023. A representative number of isolates with phenotypically detected resistance were analyzed with whole-genome sequencing. Multilocus sequence typing (MLST) was determined using genomic sequences as input data against the standard MLST schemes. High rates of resistance were detected to tetracycline (45% of C. jejuni and 26.7% of C. coli) and ciprofloxacin (81% of C. jejuni and 46.7% of C. coli), while combined ciprofloxacin/tetracycline resistance was observed in 43% of C. jejuni and 7% of C. coli isolates. The majority of whole-genome sequenced isolates possess tet(O/32/O) gene and gyrA.T861 mutation, conferring resistance to tetracycline and fluoroquinolones. All isolates have at least one resistance gene for β-lactams (blaOXA-61, blaOXA-193, blaOXA-450, blaOXA-451, blaOXA-452, blaOXA-461, and blaOXA-489), while 40% of isolates possess ant(6)-Ia for aminoglycoside resistance. In addition, a newly emerged, phenotypically identified multidrug-resistant C. coli, harbored tet(O/32/O) and tet(O) genes, point mutation in gyrA gene, and A2075G mutation in 23S rRNA. Virulence analysis highlighted C. jejuni’s broad pathogenic potential, including motility, adherence, invasion, exotoxin production, and immune modulation (e.g., wlaN gene, linked to Guillain–Barré syndrome). On the contrary, C. coli isolates possess genes exclusively for motility. Nine different C. jejuni MLST sequence types were identified, while all C. coli isolates belong to the same 828 clonal complex. Escalating AMR and a broad spectrum of virulence in Campylobacter spp. highlight the importance of continuous surveillance on the phenotypic and genotypic level, thereby allowing more efficient health care management of these infections.
Keywords
Introduction
Campylobacter enteritis, a leading cause of foodborne diarrhea globally, demonstrates variable incidence influenced by a country’s hygiene standards, with significant prevalence worldwide (Kaakoush et al., 2015). Particularly in Western nations such as the United States, Campylobacter infections have been recorded at 19.2 cases per 100,000 individuals (Delahoy et al., 2023). Alarmingly, rates are rising in North America, Europe, and Australia, while the disease remains endemic in parts of Africa, Asia, and the Middle East, especially among children (Kaakoush et al., 2015). Despite relatively low mortality, the economic burden of Campylobacter infection in the health care system is substantial (Buzby et al., 1997; Skirrow, 1991), estimated at US$2.18 billion annually in the United States (Cribb et al., 2024).
Campylobacteriosis is usually a self-limited infection, sometimes with acute and chronic complications (Kaakoush et al., 2015; Kapperud et al., 1992; Nielsen et al., 2010; Takahashi et al., 2005). Antibiotic intervention is generally not recommended, as it only marginally shortens symptom duration and may extend bacterial shedding, potentially leading to chronic carriage (Ternhag et al., 2007). However, in severe cases or among high-risk patients, azithromycin or fluoroquinolones are indicated (Ludvigsson, 2001; Mehta et al., 2017). Resistance to β-lactams and trimethoprim is common, but a recent study has shown promising susceptibility to amoxicillin–clavulanate (Schiaffino et al., 2019). The motility of Campylobacter is a critical virulence factor, driven by sophisticated flagella and a chemosensory system. Notably, recent genomic studies have uncovered that this system doubles as a Type III Secretion System, playing a crucial role in host cell invasion (Bolton, 2015).
Although there is a growing interest in researching the genetic background of the pathogenesis and antimicrobial resistance (AMR) of Campylobacter worldwide, most Croatian studies describe the epidemiology of human campylobacteriosis (Carev et al., 2017, 2018; Popović-Uroić, 1989; Viduka et al., 2024). Recently, the first molecular analysis of AMR was published (Šoprek et al., 2022). The present study aims to investigate AMR and virulence in Campylobacter jejuni and Campylobacter coli clinical isolates through whole-genome sequencing (WGS).
Materials and Methods
Setting, study design, and definitions
A retrospective observational study was conducted at the Department of Clinical Microbiology, University Hospital of Split (UHS), Croatia, from January to June 2023. A case was defined as a C. jejuni or C. coli strain isolated from diarrheal stool samples and resistant to at least one tested antibiotic during the study period. All first, unduplicated strains of C. jejuni and C. coli from stool culture were included in the study. Recurrent isolates from subsequent stool samples from the same patient were not included in the study.
Microbiological analyses
For cultivation and strain identification, all Campylobacter isolates were grown on Karmali agar (Biolife, Italy) at 42°C under microaerobic conditions (Genbox, bioMerieux, France). Suspected colonies were identified to species level by matrix-assisted laser desorption/ionisation time-of-flight mass-spectrometry (MALDI-TOF MS) (microflex® LRF, Bruker, Germany). ATCC C. jejuni 33560 was used as quality control for susceptibility testing.
Antimicrobial susceptibility testing was performed for ciprofloxacin, erythromycin, and tetracycline, by the standardized disk-diffusion method (Liofilchem, Italy) on Mueller–Hinton agar with 5% defibrinated horse blood and 20 mg/L β-NAD (MH-F) according to the EUCAST (The European Committee on Antimicrobial Susceptibility Testing, 2023) protocol (https://www.eucast.org/clinical_breakpoints). Isolates were classified as susceptible in the standard dosing regimen (S), susceptible with increased exposure (I) or resistant (R) to ciprofloxacin as well as susceptible in the standard dosing regimen or resistant to erythromycin and tetracycline, according to the EUCAST criteria. Susceptibility testing to other classes of antibiotics was not performed since there are no standardized EUCAST protocols. Multidrug resistance (MDR) was defined as acquired nonsusceptibility to at least one agent in three or more antimicrobial categories (Magiorakos et al., 2012).
Multilocus sequence typing and WGS
Fifteen representative isolates (3 strains of C. coli and 12 strains of C. jejuni) were selected for WGS based on phenotypically detected AMR. DNA from bacterial isolates was extracted with the NucleoSpin Tissue kit (Macherey-Nagel, Germany) according to the manufacturer’s protocol and 3-h proteinase K digestion at 56°C. Extracted DNA was quantified based on fluorescence with a Qubit 4 Fluorometer using a Qubit 1X dsDNA HS Assay Kit (Invitrogen, USA). DNA was normalized using the Myra liquid handling system (BMS, Australia) to 3.3 ng/µL. The WGS library was prepared with the Bravo Option B liquid handling system (Agilent, USA), using the Illumina DNA prep kit, and sequenced on an Illumina NextSeq 550 instrument (Illumina, USA).
De novo assembly was performed with the SPAdes genome assembler v3.10.1. (Prjibelski et al., 2020). The obtained FASTA files were analyzed using ResFinder 4.1 (Bortolaia et al., 2020; Camacho et al., 2009; Zankari et al., 2017). The minimal percentage of identity between the best-matching resistance gene in the database and the corresponding sequence in the genome was set to 90, and the minimum length of coverage that a sequence must overlap a resistance gene to count as a hit for that gene was set to 60%.
The sequences of seven housekeeping genes in the genome were used to determine the sequence type (ST) of C. jejuni and C. coli isolates through comparison with PubMLST (PubMLST—Public Databases for Molecular Typing and Microbial Genome Diversity Available online: https://pubmlst.org/). The sequences included in the study were registered with the project database https://www.ebi.ac.uk/ena/browser/view/PRJEB64612 (Wu et al., 2023).
Statistical analysis
Fisher’s exact test was applied to evaluate differences in antibiotic resistance phenotypes and the presence of specific resistance genes between C. coli and C. jejuni samples. A p value of less than 0.05 was considered statistically significant.
Results
C. coli and C. jejuni phenotypic resistance profile
Phenotypical resistance of all 115 isolates is shown in Table 1. C. coli showed full resistance to ciprofloxacin in 7 out of 15 isolates (46.6%) and intermediate resistance in 8 out of 15 isolates (53.3%), whereas C. jejuni exhibited a higher full resistance rate of 81 out of 100 isolates (81%). For tetracycline, C. coli exhibited resistance in 4 out of 15 isolates (26.7%), while C. jejuni showed resistance in 45 out of 100 isolates (45%). All C. jejuni isolates were susceptible to erythromycin, contrasting with one resistant C. coli isolate.
Antimicrobial Susceptibility Patterns of Campylobacter coli and Campylobacter jejuni Isolates

Fisher’s exact test showed significant difference for ciprofloxacin resistance.
Fisher’s exact test for ciprofloxacin resistance showed a significant difference between the C. coli and C. jejuni. In contrast, the observed differences in tetracycline and erythromycin resistance are not statistically significant. The coresistance to ciprofloxacin and tetracycline was high in both Campylobacter groups: C. coli (26.8%) and C. jejuni (43%).
WGS analysis
Multilocus sequence typing and distribution of antimicrobial resistant genes
The identified multilocus sequence typing (MLST) STs and clonal complexes (CCs) are presented in Table 2. We identified nine different MLST STs and six different MLST CCs. The most predominant C. jejuni were ST6461 (CC-353) and ST122 (CC-206). The remaining C. jejuni isolates belong to ST50 (CC-21), ST367 (CC-257), ST464 (CC-464), and ST9897 (one isolate each). All C. coli isolates belong to the same 828 CC.
Multilocus Sequence Typing Characterization of Campylobacter Strains
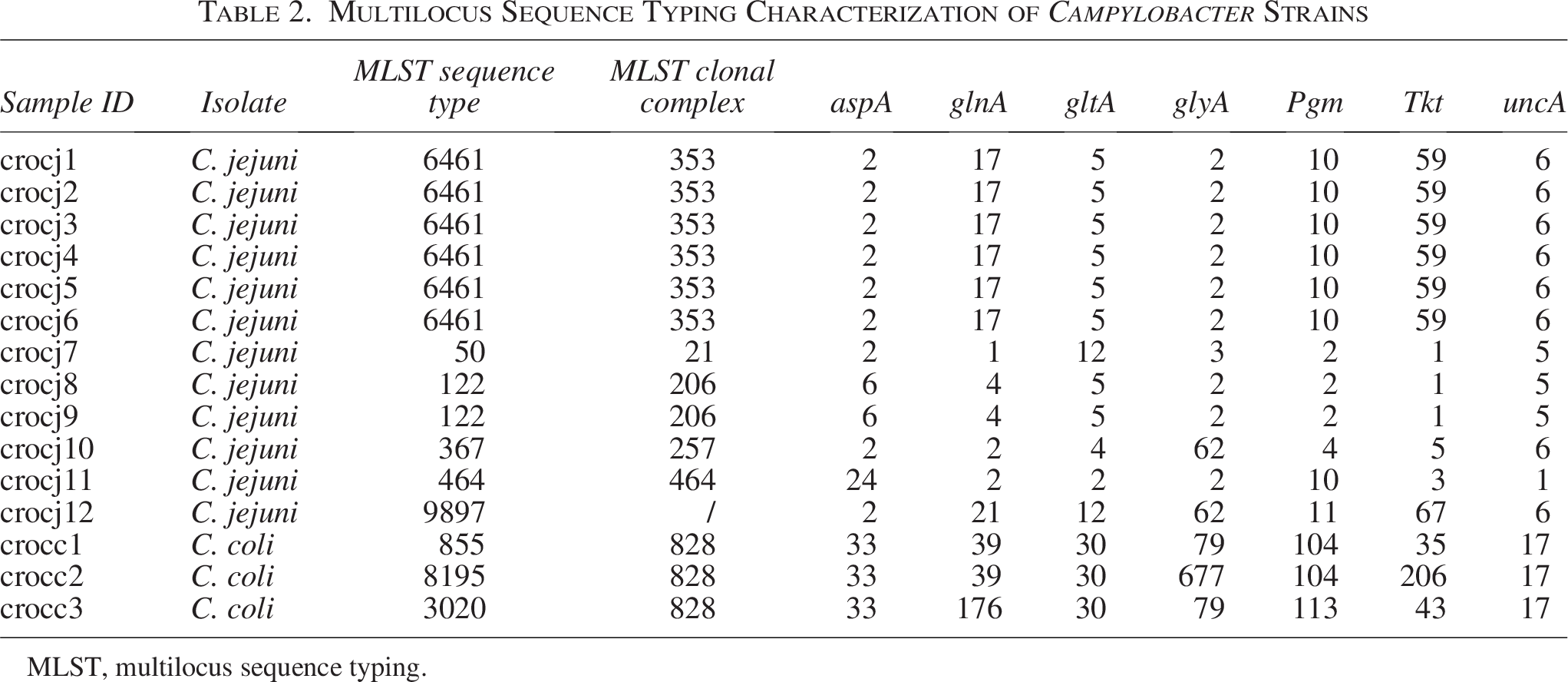
MLST, multilocus sequence typing.
A detailed analysis of the distribution of AMR genes from all 15 Campylobacter isolates is shown in Table 3. The genetic basis for fluoroquinolone resistance, T861 point mutation in gyrA, was detected in most of the tested isolates (14/15, 93.3%). WGS revealed the presence of the tet(O/32/O) gene in 14 out of 15 isolates (93.3%), while four isolates (26.7%) additionally harbored the tet(O) gene, both associated with tetracycline resistance. Macrolide resistance was observed in one isolate (1/15, 6.7%) and was associated with the 23S rRNA 2075A>G gene mutation. Aminoglycoside resistance, linked to the ant(6)-Ia gene mutation, was found in six out of 15 isolates (40%), all within the C. jejuni family, while aph(3′)-III was detected in one C. coli isolate (6.7%). All isolates had at least one gene for resistance toward β-lactam antibiotics.
Distribution of Antimicrobial Resistance Genes in Campylobacter Strains Detected with Whole-Genome Sequencing
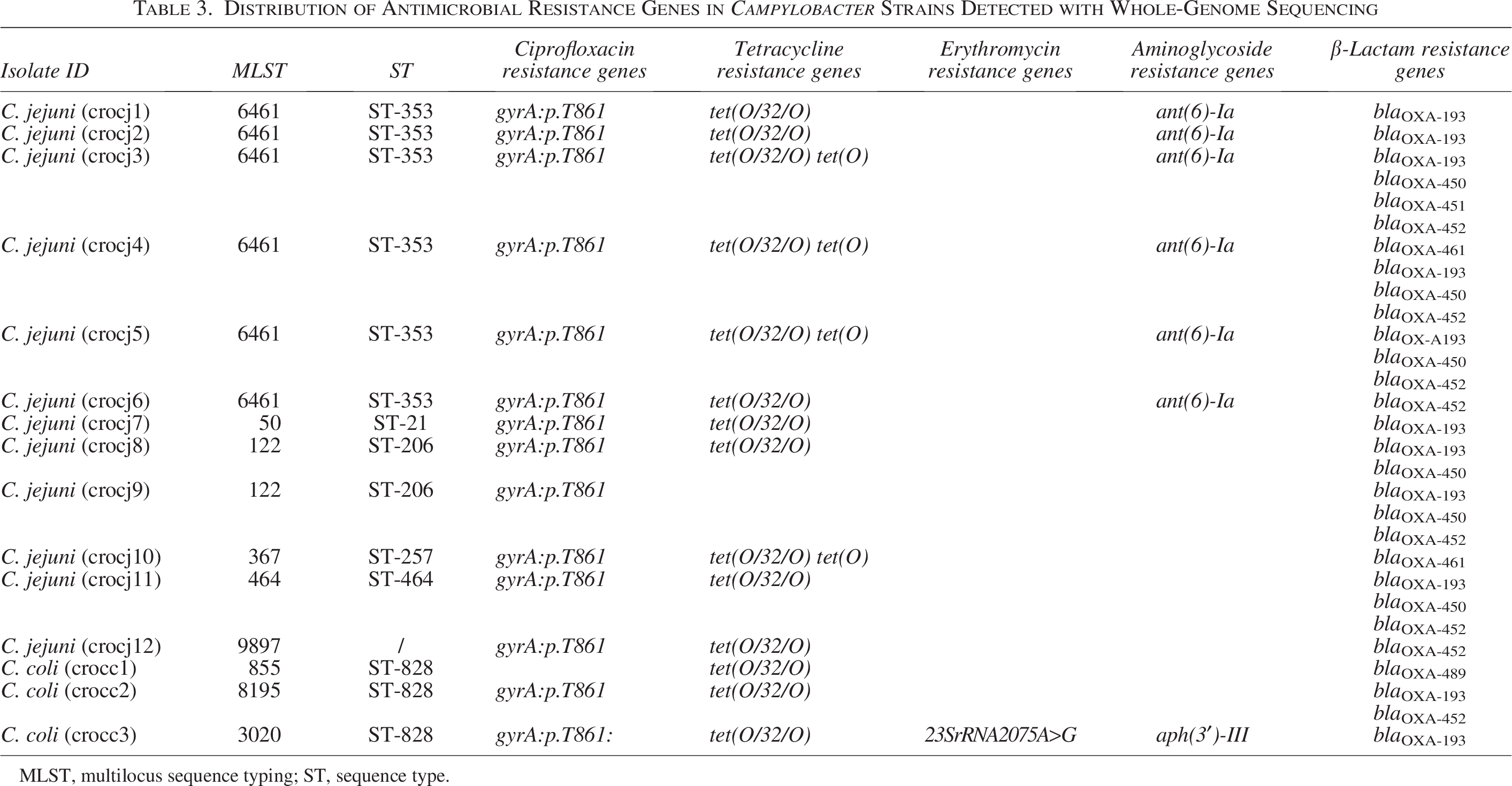
MLST, multilocus sequence typing; ST, sequence type.
A newly identified multidrug-resistant C. coli strain, phenotypically resistant to ciprofloxacin, tetracycline, and erythromycin, harbored a variety of resistance genes: tet(O/32/O), tet(O), blaOXA-193, and aph(3′)-III, as well as gyrA.T861 and 23S rRNA 2075A>G gene mutations.
Grouping of resistance genes within the same CC was observed. More precisely, an almost identical resistotype was detected for all isolates belonging to MLST 6461, CC-353 (gyrA.T861, tet(O/32/O), ant(6)-Ia, blaOXA-193), with the only difference being that some isolates carried additional blaOXA genes.
In addition, the presence of resistance genes was compared between C. coli and C. jejuni isolates, with no statistically significant differences between these two groups (Table 4). The gyrA.T861 mutation was found in 66.7% of C. coli isolates and in all C. jejuni isolates, while the tet(O/32/O) gene was present in all C. coli isolates and in most of the C. jejuni isolates (91.7%). The additional tetracycline resistance gene, tet(O), was found only in C. jejuni strains (33.3%).
Resistance Genes and Associated Phenotypes in Campylobacter coli and Campylobacter jejuni Isolates
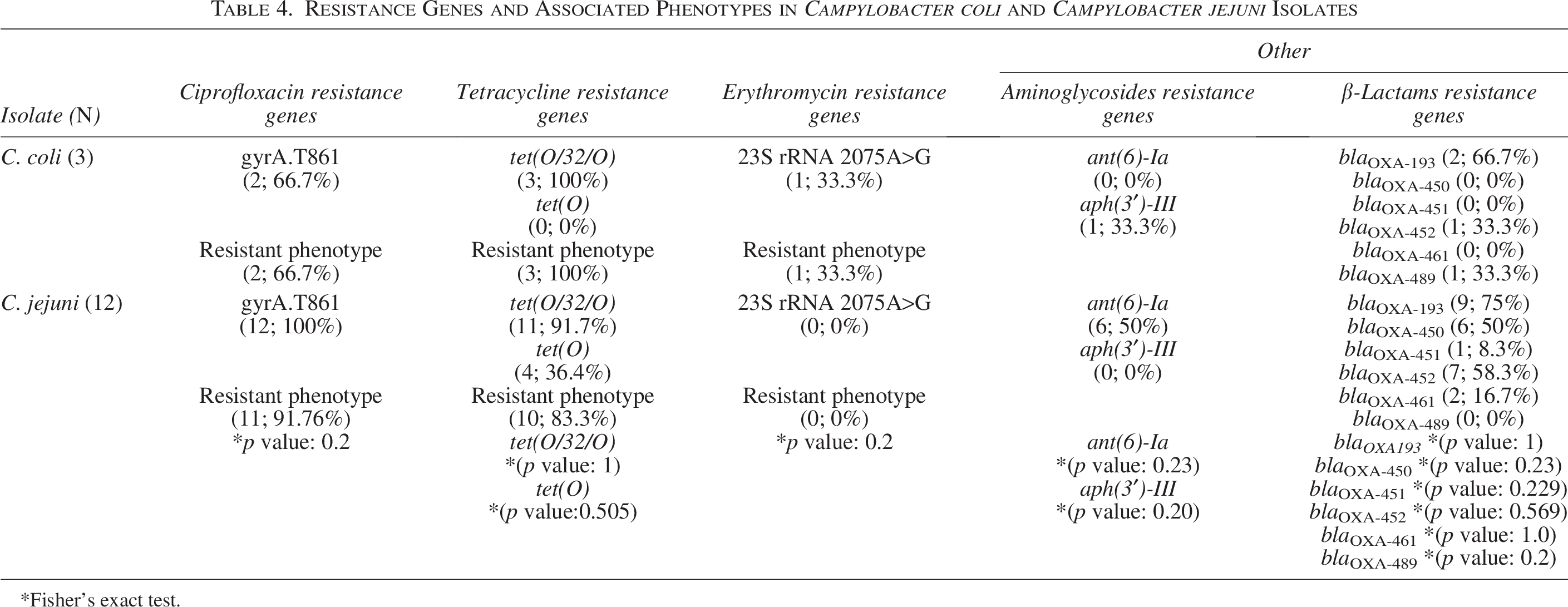
Fisher’s exact test.
Distribution of Virulence Genes in Campylobacter Strains
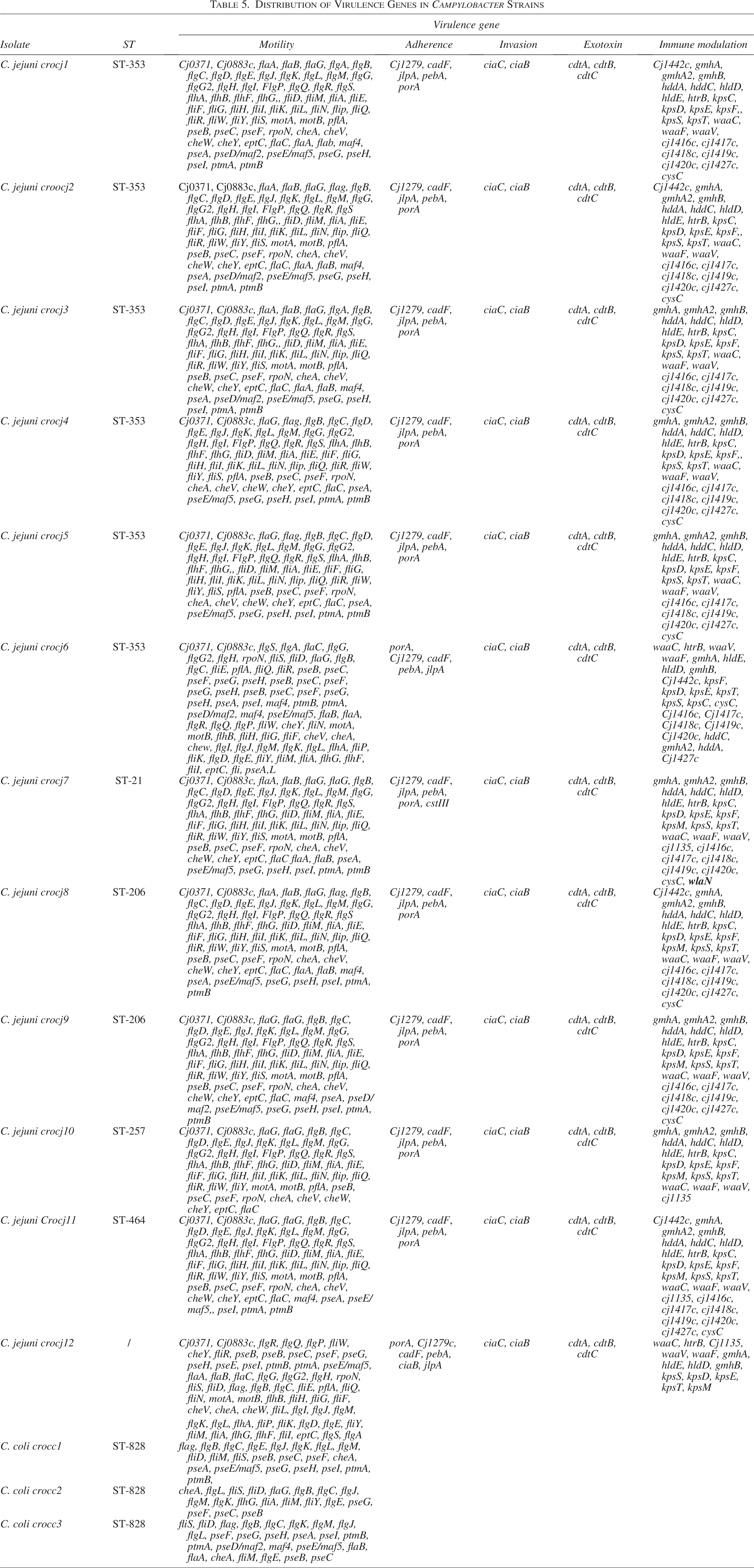
Regarding the distribution of aminoglycoside resistance genes, the ant(6)-Ia gene was not detected in any of the C. coli isolates but was present in 50% of the C. jejuni isolates. On the contrary, the aph(3′)-III gene was detected only in one C. coli strain.
For β-lactam resistance genes, C. coli isolates showed the following distribution: blaOXA-193, blaOXA -452 , and blaOXA-489. In comparison, C. jejuni isolates exhibited the following distribution: blaOXA-193, blaOXA-450, blaOXA-451, blaOXA-452, and blaOXA-461.
The only A2075G point mutation in 23S rRNA identified in this study was found in a C. coli isolate.
Distribution of virulence genes
Detailed analysis of virulence genes is shown in Table 5. All C. jejuni isolates possess multiple virulence genes for motility, adherence (e.g., the cadF gene for outer membrane fibronectin-binding protein synthesis, and the porA gene for major outer membrane protein production), invasion (ciaC and ciaB), exotoxin production (cdtA, cdtB, and cdtC), and immunomodulation (Cj1442c, gmhA, gmhA2). In contrast, C. coli strains have virulence genes exclusively for motility, as shown in Table 5.
For both strains, the most numerous genes were those related to motility, some of which code for flagella synthesis (e.g., fliQ), flagellin (flaA), flagellar hook-basal body complex proteins (e.g., fliE), flagellar motor proteins (motA and motB), chemotaxis proteins (e.g., cheV and cheW), and flagellar rod assembly proteins/muramidases (flgJ).
Regarding immunomodulation, a variety of genes were identified in C. jejuni strains, including different kps and Cj1416 genes coding for various capsule polysaccharide proteins, and waaV, waaC, and Cj1135 genes for lipooligosaccharide production. Notably, one of the most significant genes, wlaN, responsible for the Guillain–Barré syndrome, was found in a C. jejuni isolate.
Discussion
There is a growing need to find and implement accurate and rapid routine laboratory methods that can quickly detect AMR in Campylobacter isolates from clinical specimens and identify patients who might be infected with more virulent strains associated with worsening clinical outcomes (Gharbi et al., 2022).
Up to now, there are more than 39,700 C. jejuni and 11,900 C. coli genomes available in public repositories. Total global analysis determined that resistance to β-lactams and tetracycline was the most frequent for both species, with resistance to quinolones as the third-most important in C. jejuni and to aminoglycosides in C. coli (Cobo-Díaz et al., 2021).
The present study has unveiled high levels of resistance in Campylobacter spp. to quinolones and tetracyclines. When considered together, all Campylobacter strains show some level of quinolone resistance, but C. coli seemed to exhibit a weaker level of resistance compared with C. jejuni, which is consistent with previous findings (Cobo-Díaz et al., 2021).
C. jejuni strains were 81% resistant to quinolones, while the remaining 19% of strains were susceptible with increased antibiotic exposure. These figures represent a substantial rise compared with past national statistics, where the Croatian resistance rate in 2020 was 71% (Šoprek et al., 2022). Fluoroquinolone resistance in C. jejuni in the Split-Dalmatia County has slowly increased from 60% in 2013 (Carev et al., 2017) to 74.8% in 2021 (Viduka et al., 2024). Also, ciprofloxacin/tetracycline coresistance is on the rise, from 21.3% in 2013 to 43% in the present study (Carev et al., 2017).
When analyzing current data from the European Food Safety Authority (EFSA)/ECDC, C. jejuni resistance to ciprofloxacin found in this study aligns more closely with the southern European countries, where resistance rates also fall between 80% and 100%. This contrasts with the rest of the European Union, where the average resistance is approximately 61.2%, and Western Europe, where it hovers around 60% (Antimicrobial Resistance in Europe Available online at: https://multimedia.efsa.eu/dataviz-2020/index.htm).
Resistance to ciprofloxacin in C. coli in our study reflects a broader trend observed across the European Union, where the resistance rate is 65.8%. In contrast, Western Europe (including Austria, France, and the Netherlands) reports resistance rates ranging from 50% to 70%. Southern Europe, particularly countries such as Italy, Portugal, and Malta, shows even higher resistance levels, reaching 80% to 100% (Antimicrobial Resistance in Europe Available online at: https://multimedia.efsa.europa.eu/dataviz-2020/index.htm) (EFSA). Notably, Split’s resistance rates are slightly lower than these southern counterparts.
Regarding tetracycline resistance, C. coli in Croatia shows a markedly lower resistance rate of 26.7%, compared with the European Union’s average of 74%. This pattern of lower resistance in Croatia resembles that of Eastern Europe, where resistance varies widely from 20% to 50%, unlike the uniformly high rates (80–100%) observed in both Western and Eastern Europe. Tetracycline resistance in C. jejuni in Split is approximately 45%, closely mirroring the European Union’s average of 43.7%. This suggests a consistent resistance profile across different regions of Europe for this antibiotic (Antimicrobial Resistance in Europe Available online at: https://multimedia.efsa.europa.eu/dataviz-2020/index.htm).
Erythromycin resistance in C. jejuni is not detected in our study, which is similar to the European Union’s average of 0.7%. Poland, however, stands out with a resistance rate of 10%. For C. coli, erythromycin resistance (coded with the 23S rRNA 2075A>G gene mutation) is newly emerging in Split, although it has already reached around 10% across the European Union. This resistance does not follow a clear regional pattern within Europe; for instance, Portugal, the Netherlands, and Finland report rates of 36.8%, 20.4%, and 43%, respectively, while Austria, Slovenia, and Slovakia have significantly lower rates (Dahl et al., 2021). In Central China, the resistance is around 25.2%, predominantly given the 23S rRNA.2075A>G, which is commonly copresent with the erm(B) gene. Clinical isolates from Beijing, however, show varied resistance mechanisms, including mutations in the 50S rRNA_L22_A103V and the cmeABC operon, highlighting regional differences in resistance patterns (Cheng et al., 2020; Zhang et al., 2023). For this reason, the occurrence and the first molecular characterization of macrolide-resistant C. coli Croatian clinical isolate are extremely important for future national and global monitoring and for the control of the development and spread of resistance.
All strains analyzed in this study possess various blaOXA genes, which encode resistance to different β-lactams. Moreover, a previous study showed that resistance to ampicillin and ceftriaxone is more than 40–50%, which predicts clinical failure if used in empirical therapy (Schiaffino et al., 2019). When looking at aminoglycoside resistance genes, in our study, C. coli seems to have solely aph(3′)-III and C. jejuni ant(6)-Ia genes, showing a different gene pattern for developing resistance.
In addition, it is important to mention that Campylobacter, as a foodborne pathogen, may possess different AMR patterns, not only in different countries but also in different hosts. The Croatian study highlighted a lower resistance in seagull strains compared with human and poultry strains (Jurinović et al., 2020). In China, the fexA gene (conferring resistance to florfenicol), frequently found within tet(L)-fexA-tet(O) MDR genomic islands of Campylobacter isolated from poultry and pigs, was not detected in our study (Tang B et al., 2020; Tang B et al.., 2021; Tang B et al.., 2022).
Regarding the virulence genes, WGS analysis revealed different virulence patterns between C. coli and C. jejuni, where C. coli had exclusively genes for bacterial motility, while C. jejuni had numerous virulence genes for bacterial adherence, invasion, toxin production, and immunomodulation. According to significant differences in virulence gene composition, C. jejuni may have a predisposition to cause more severe clinical outcomes compared with C. coli. However, only a limited number of C. coli isolates were tested with WGS and this hypothesis should be tested with the larger sample size. On the contrary, motility is one of the most important mechanisms in the pathogenesis of Campylobacter infections, so the lack of other virulence genes does not exclude an unfavorable outcome of acute infection.
The presence of the LOSSIAL-related (sialylated lipooligosaccharide) gene, wlaN, was detected in a C. jejuni strain for the first time in Croatia. This gene can trigger the Guillain–Barré syndrome by coding for the β-1,3-galactosyltransferase enzyme that is required for the production of LOSSIAL. In our study, the wlaN gene was found in an isolate belonging to the MLST 21 CC, which was also shown by Guirado et al.. in their study (on 150 isolates) (Guirado et al., 2020).
Geographical differences in CC distribution were found for C. jejuni and in resistome genotypes for both C. jejuni and C. coli species (Cobo-Díaz et al., 2021). In the present study, there were nine different MLST STs, with ST6461 (CC-353) and ST122 (CC-206) being the most predominant C. jejuni, while all C. coli isolates belong to the same 828 CC.
Various STs detected in the present study have a worldwide distribution. C. jejuni MLST ST 6461 (CC-353) is also the most common CC reported from Eastern China (Zhang et al., 2015), while C. jejuni belonging to MLST STs 21 and 257 was described previously by Sarhangi et al.. as the most predominant CC in Tehran (Sarhangi et al., 2021). Several STs detected in this study (464, 257, 6461, 21, and 122) have been reported from Poland (Wieczorek et al., 2020).
Even though there is no strong evidence about the connection of certain STs, virulence, and resistance, all isolates belonging to ST 6461 have almost identical resistotype, while C. jejuni isolate belonging to MLST 21 exhibits a robust array of virulence factors, including the wlaN gene, suggesting a potential for increased virulence and severe infection outcome.
Conclusion
The study underscores the escalating AMR to ciprofloxacin and tetracycline in Campylobacter spp, with emerging resistance to erythromycin in C. coli isolated in Croatia, highlighting the importance of continuous surveillance and genomic analysis. These resistance patterns reflect broader regional and global trends, necessitating urgent attention to antimicrobial stewardship and tailored treatment approaches.
Authors’ Contributions
Conceptualization: L.K., A.N., and G.O. Data curation, formal analysis, and methodology: L.K. and A.N. Collecting data: I.F. and A.G.-G. Laboratory analysis: L.K., A.N., I.F., A.G.-G., I.G.-B., Z.R., M.R.-S., J.M., M.T., D.S., J.K., L.Š.A., I.T., and D.J. Software: D.S., J.K., L.Š.A., and D.J. Funding acquisition: I.F., I.T., and A.N. Writing—original draft: L.K. and A.N. Writing—review and editing: all authors. All authors have read and agreed to the published version of the article.
Footnotes
Author Disclosure Statement
The authors have no relevant financial or nonfinancial interests to disclose.
Funding Information
This research was conducted as a part of the implementation of WGS for detection of AMR under the HERA2 project, which received funding from the European Union’s Health and Digital Executive Agency (HaDEA) under Grant Agreement No 101102440. The authors gratefully acknowledge additional funding provided by The ESCMID Study Group for Food- and Waterborne Infections (EFWISG).