
Abstract
The study of the human nervous system remains challenging due to its inherent complexity and difficulty in obtaining original samples. Three-dimensional (3D) bioprinting is a rapidly evolving technology in the field of tissue engineering that has made significant contributions to several disciplines, including neuroscience. In order to more accurately reflect the intricate multicellular milieu of the in vivo environment, an increasing number of studies have commenced experimentation with the coprinting of diverse cell types. This article provides an overview of technical details and the application of 3D bioprinting with multiple cell types in the field of neuroscience, focusing on the challenges of coprinting and the research conducted based on multicellular printing. This review discusses cell interactions in coprinting systems, stem cell applications, the construction of brain-like organoids, the establishment of disease models, and the potential for integrating 3D bioprinting with other 3D culture techniques.
Impact Statement
In the field of neuroscience, 3D bioprinting is becoming a key tool for creating neural tissues. These techniques aim to mimic the complex cellular environments found in the body. Recent advancements in neural models using multicellular bioprinting show the importance of optimizing printing parameters. This optimization enhances cell survival and tissue functionality. Studies also focus on how different types of neural cells interact within these printed structures. Despite these advances, there is a lack of comprehensive reviews on using multicellular bioprinting for neural tissues. This article aims to fill that gap. It summarizes the challenges and progress in multicellular neural bioprinting. It helps researchers choose the best printing parameters and model construction methods for neural tissue engineering. This review also explores the current status and limitations of multicellular bioprinting in neural tissues. It supports further innovation in this area. The findings will aid in advancing neural bioprinting technologies and improving future neural tissue designs.
Introduction
Brain research has remained robust, driven by curiosity about human nature and the persistent challenges of brain-related diseases. Long before Gauguin’s inquiries, humans had started the pursuit of a deeper understanding about themselves, intensified by technological advancements. Ethical concerns restrict the use of human brain tissue while species differences limit the accuracy of animal models. Consequently, developing more accurate in vitro human neural models has become a key focus for researchers.
In vivo brain models include two-dimensional (2D) cell models, 3D cell models (spheroids, organoids, 3D bioprinting), and embedded animal models. Since the native neural tissue is a 3D structure primarily composed of neurons, glial cells, and other cell types arranged in a highly ordered manner, it is crucial to consider the interplay between these cell types and their spatial organization when constructing in vitro neural models. 1 Compared with other tissue engineering platforms, 3D bioprinting offers distinct advantages, including the capacity to print multilayered or complex structures, dictate multicellular distribution, and adjust local environments to support the growth of diverse cell types.2–5 These features are particularly advantageous for the construction of neural models with multiple structural layers and cell types in vitro. 6 In recent years, advances have been made in printing specific types of nerve cell, paving the way for the development of multicellular neural tissue bioprinting (Fig. 1).7–9

Pipeline design for neural 3D-bioprinting with multiple cell types. Common types of cells used in neural biofabrication include neurons, glia cells, and stem cells (left). Bioink loaded with viable cells are then subjected to bioprinting to yield 3D constructs that mimic in vivo neural tissues (middle). Establishing multicellular 3D-printed neural models can facilitate neurophysiological, neuropathological, and mechanistic studies, as well as drug screening in vitro (right).
Multicellular Neural Tissue Bioprinting
Ensuring the postprint viability of neural cells remains a fundamental challenge in 3D bioprinting, a problem that has become increasingly pronounced as printing systems grow increasingly intricate. In multicellular neural tissue printing, concerns extend beyond shear stress-induced damage and include issues in bioinks, cross-linking, and microenvironmental factors.10–12 Stem cells have been extensively employed in bioprinting applications. Given their sensitivity to environmental parameters, ensuring cell viability, pluripotency, and controlled differentiation both during and after the printing process is of significant interest. 13
In this section, we summarize the current research and development aimed at enhancing multicellular viability in neural tissue printing. This goal is being achieved from three perspectives: optimization of printing protocols, selection of bioinks, and biomaterial scaffolds. In addition, we discuss the various stem cells used in neurobioprinting research, as well as their potential applications and related challenges.
Printing protocols
The prevailing bioprinting configurations can be broadly classified into three categories: extrusion-based, inkjet-based, and laser-assisted techniques. 14 When engaging in multicellular neural tissue printing, a primary concern lies in addressing the damage incurred by shear forces during the printing process. Nozzle-based methods are significantly impacted by shear forces, whereas laser-assisted techniques demonstrate superior performance in terms of cell viability and printing precision (Figure 2).15–18
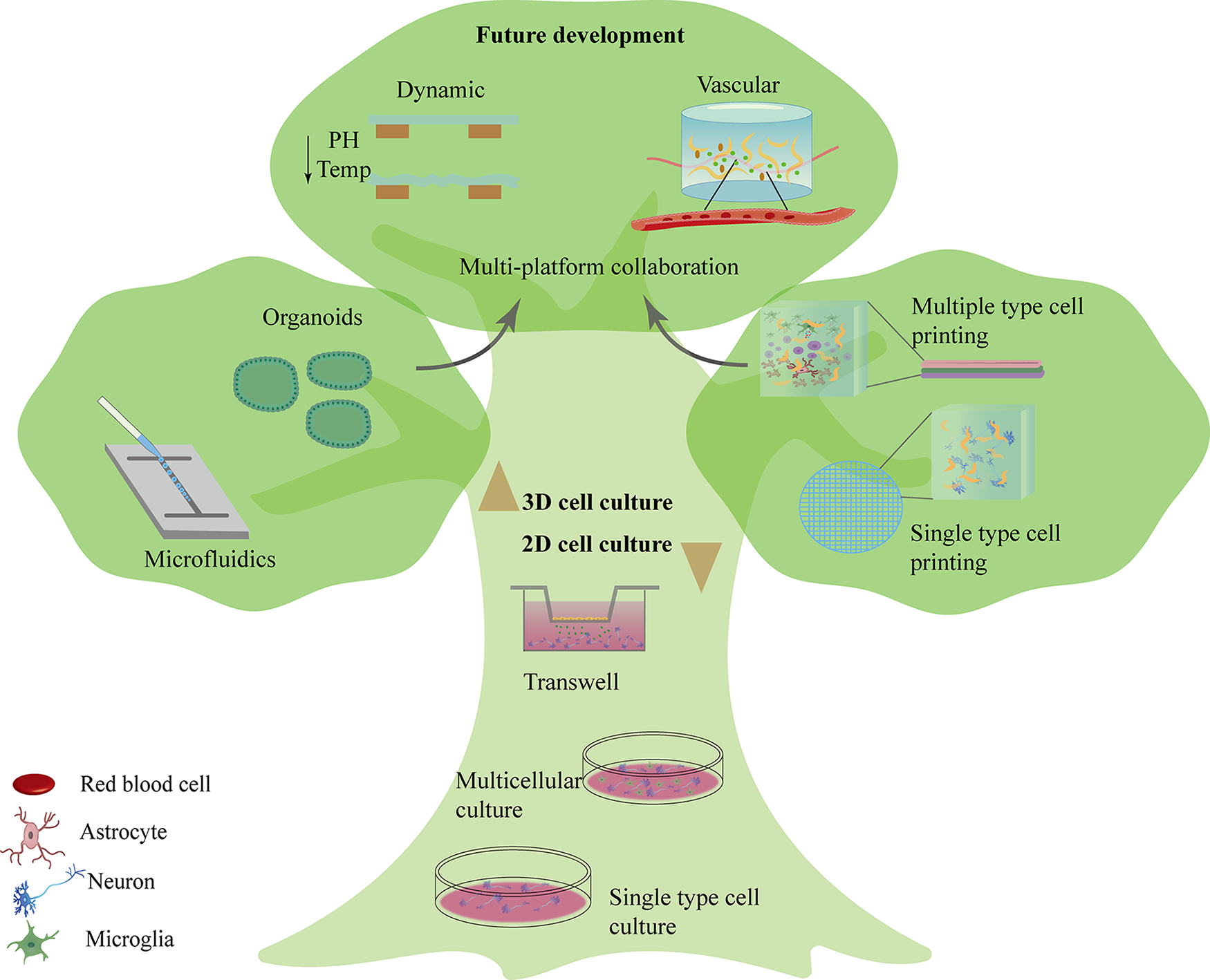
The evolution and future prospects of in vitro neural models. Initial approaches advanced from neuronal cell culture of single-cell types to coculture of multiple neuronal cell types (bottom). 3D culture techniques then emerged, including self-assembled organoids, microfluidics2–5 (left), as well as 3D bioprinting1,19–22 (right) that underwent a progression from single to multiple cell types.20,23–26 Future prospects for 3D bioprinting-based in vitro neural models (top) include dynamic bioinks,27–29 combined use with other 3D culture platforms,16,30–35 and development of vascular systems.16–18
Since distinct optimal printing windows have been described for different cell types, it is of the utmost importance to identify the equilibrium point for simultaneously printing various cells (see Table 1). In the case of Schwann cells (SCs, see Table 4 for all abbreviations) and mesenchymal stem cells (MSCs), maintaining the injection pressure within specific ranges (50 kPa for cylindrical nozzles; 10 kPa for conical nozzles) can effectively protect both cell types from shear force-induced injury. 19 Neurons and astrocytes demonstrate better resilience during printing under a printing pressure of 150 kPa, retaining excellent neural network formation capacity in subsequent 3D coculture systems of neurons and astrocytes. 20
Nerve Cell Printing Parameters

ALG, alginate; BC, border cap neural crest stem cells; CNF, cellulose nanofibril; GEL, gelatin; GelMA, gelatin methacrylate; HA, hyaluronic acid; HAMA, hyaluronic acid methacrylate; hiPSCs, human induced pluripotent stem cell; hNPC, human nucleus pulposus cells; MN, motor neuron; MnTBAP, manganese III tetrakis 4-benzoic acid porphyrin; mTG, microtransglutaminase; NSC, neural stem cells; SA, sodium alginate; UV, ultraviolet.
The use of ultraviolet (UV) irradiation for cross-linking is a common strategy in printing.21,22 Given the thicker hydrogel structures commonly encountered in brain-like constructs, particularly in multicellular systems where layer-by-layer printing is frequently employed, extended durations and higher intensities of UV irradiation are often required in multicellular neural tissue printing. This requires encapsulated neural cells to be exposed to higher-intensity UV radiation to maintain structural stability, while prolonged UV exposure inevitably results in multicellular damage. 44 Multicellular to lerance to UV radiation varies, with astrocytes demonstrating the capacity to maintain 80% viability following 100 s of exposure at an intensity of 800 mW. 45 In comparison, neurons are relatively more susceptible, necessitating the implementation of a gentler strategy, for instance, 30 s at 20 mW for cross-linking. 20 It is also possible to exploit the differential cross-linking times without modifying the bioink composition by adopting an embedded printing approach. This approach entails initially printing and cross-linking the more resilient cells onto the scaffold, followed by printing less tolerant cells. 45
In addition to the optimization and refinement of existing printing protocols and parameters, the development and application of new techniques are also underway. A number of new approaches have been identified that are already being applied in multicellular printing or may be applied in the future (see Table 2).
Printing Process and Bioink Optimization
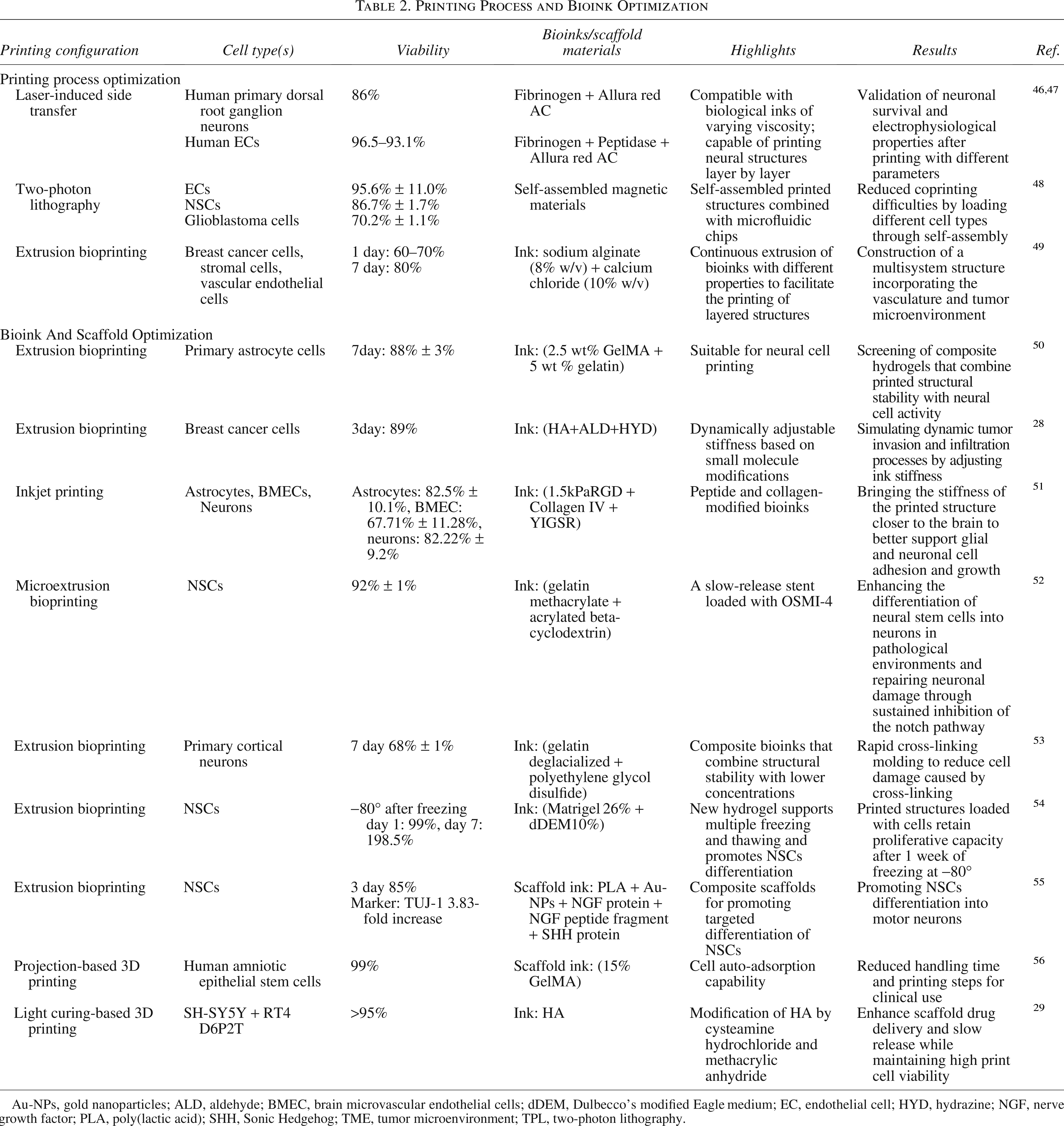
Au-NPs, gold nanoparticles; ALD, aldehyde; BMEC, brain microvascular endothelial cells; dDEM, Dulbecco’s modified Eagle medium; EC, endothelial cell; HYD, hydrazine; NGF, nerve growth factor; PLA, poly(lactic acid); SHH, Sonic Hedgehog; TME, tumor microenvironment; TPL, two-photon lithography.
Bioink
Commonly employed bioinks in conventional bioprinting include gelatin (GEL), hyaluronic acid (HA), and poly(lactic acid).41,57 Recently, with the increasing recognition of the importance of the extracellular matrix (ECM), several studies have been focusing on utilizing ECM directly as bioinks in the printing process.37,58,59
When conducting bioprinting of multiphasic neural cells, it is essential to accommodate the optimal survival environments for diverse cell types, particularly considering the heightened fragility of neural cells compared with other cell types. For instance, under identical printing conditions (gelatin methacrylate [GelMa]), fibroblasts (FIB) exhibit significantly higher viability than neuronal cells (FIB viability: >90%; neuronal cell viability: 20%). 9
Ensuring high viability for different cell types within the final printed construct is therefore a fundamental aspect of multicellular neural tissue bioprinting. Sullivan et al. were able to simultaneously support the growth requirements of several different neuronal cell types on a single substrate by incorporating adhesive peptides and protein (e.g., fibronectin or laminin) into the bioink, resulting in viability rates of (astrocyte viability: 82% ± 10.1%, endothelial cells [EC] viability: 67.71% ± 11.28%, neuronal cell viability: 82.22% ± 9.2%). 51 The development and application of particulate hydrogels provide another approach. In comparison to their bulk counterparts, particulate hydrogels demonstrate enhanced efficiency in the transport of nutrients and the exchange of intercellular signaling factors, which is of paramount importance for the interactions between cells within a 3D coculture system. 60 To this end, Xu et al. developed a microparticulate hydrogel with the capacity to encapsulate neural cells. 61 In a 3D coculture system of neurons and astrocytes, the particulate hydrogel demonstrated superior compatibility with neuronal cells, as evidenced by higher printing viability (particulate hydrogel: 63% ± 7%; bulk hydrogel: 36% ± 6%) and longer neurite lengths (particulate hydrogel: 65.52 ± 11.59 μm; bulk hydrogel: 22.90 ± 4.70 μm).
When printing multicellular neural tissue, it is also essential to consider the stiffness of the bioink.62,63 The brain is softer than many other tissues (with a stiffness of 0.1–1 kPa); therefore, bioinks with lower stiffness help maintain cell survival after printing. 29 Adding layers can help recreate the natural layering of neural tissue, but at the meantime affects the stiffness. A better choice may be composite bioinks that balance how they are printed and how stiff they are (see Table 2). 50 It is worth noting that although a lower stiffness is typically regarded as advantageous for printing neural cells, the specific parameters must be adapted to align with the experimental objectives. For example, when modeling brain aging, the increased stiffness in certain brain regions due to aging must be taken into account 64 ; when studying oligodendrocytes and myelin-related content, the stiffness requirements during myelination must be considered.65,66 A hydrogel with the ability to dynamically adjust its stiffness after printing can be used in complex environments. 27 Hull et al. addressed the issue of hydrogel hydrazone cross-linking by incorporating hydrazine (HYD) catalysts (aldehydes) and competitors (HYDs) into HA, thus creating a bioink with tunable stiffness postprinting. This approach permits the accommodation of varying stiffness demands during the different stages of neural printing. 28 One method of modifying the printing material is through alteration of its composition. For instance, Buckley et al. employed chemical modification of HA through thiolation and methacrylate ester, which enhanced its compatibility with neurons and glial cells while also improving the material’s capacity for drug delivery. 29
Stem cell in multicellular bioprinting
As a result of their capacity for proliferation and differentiation in vitro, stem cells are the subject of considerable interest across a number of fields, including 3D neural bioprinting.39,67–72
Induced pluripotent stem cells (iPSCs), which possess the same genetic identity as the donor, facilitate the creation of disease models that are tailored to the specific genetic characteristics of individual patients. 73 A common strategy in multicellular neural tissue bioprinting is the induced differentiation of iPSCs into different cell types. For example, Salaris et al. achieved simple 3D coculture by printing pretreated iPSCs and observing iPSCs-differentiated neuronal cells and astrocytes in the printed structures. 74 Similarly, Chloe et al. utilized iPSCs for printing and subsequently differentiated them into glutamatergic neurons and astrocytes. 75 Additionally, Sullivan et al. achieved simple 3D coculture by printing iPSCs and subsequently differentiating them into astrocytes, brain microvascular endothelial-like cells, neural progenitor cells, and neurons. 51 In multicellular bioprinting, the differentiation of stem cells is influenced not only by various printing parameters76–80 and external stimuli81–83 (see Table 1), such as lasers or pulsed ultrasound, but also by the impact of other cells within the system.
Neural stem cells (NSCs) are stem cells with the potential to differentiate between neuronal cells (70–80% glutamatergic neurons, 20–30% gamma-aminobutyric acid [GABA]-ergic neurons) and glial cells.84,85 Previous studies have demonstrated that the 3D coculture of NSCs with MSCs effectively promotes NSCs differentiation toward a neuronal lineage.86,87 In a recent study, Zhou et al. reaffirmed this phenomenon by developing a GelMA-based hydrogel scaffold capable of encapsulating both cell types. 88 The utilization of this 3D coculture platform revealed that MSCs facilitated NSCs differentiation into neurons and eventually achieved a greater reduction in spinal cord injury (SCI) lesion volume (SCI group lesion volume: 82.42% ± 1.59%; GelMA/bone marrow-derived MSCs/NSCs group lesion volume: 30.96% ± 1.02%).
In multicellular neural tissue bioprinting, it is imperative to emphasize the differentiation of stem cells postprinting. 3D coculture environment strongly influences the differentiation trajectory of stem cells. Evidence suggests that astrocytes can facilitate the differentiation of stem cells into neurons in 3D coculture systems.45,89 Moreover, astrocytes are responsible for maintaining glutamate homeostasis within the ECM through the glutamate–glutamine cycle. 45 This process sustains neuronal activity, which is beneficial for the assembly of neural networks.
The printing of stem cells following predifferentiation treatment is a prevalent method for the construction of neural tissue. 90 Common predifferentiation methods include: growth factor induction, physical stimulus induction, gene editing induction, and small molecule induction.91–93 Before performing stem cell bioprinting, the impact of predifferentiation on the postprinting survival rate needs to be considered. Although a longer predifferentiation treatment may consolidate the directed differentiation of stem cells after printing, it may simultaneously decrease the survival rate of the printed cells (pretreated for 5 days: 93.0% ± 0.6%–pretreated for 20 days: 68.4% ± 1.3%). 94 After bioprinting, it is also important to consider the influence of the culture environment on the subsequent differentiation of the stem cells. For example, even if NSCs are printed and then cultured in a medium that promotes differentiation into neurons, the NSCs may still differentiate into glial cells. 95
In multicellular bioprinting, there are many other factors that affect the differentiation of stem cells into nerve cells, such as the culture environment after printing (Fig. 3). It has been demonstrated that if the stem cells are in a pathological environment this may lead to a greater tendency to differentiate toward glial cells rather than neurons. 52 In addition, modifying the scaffolds,96–98 preparing specific bioinks,76,99,100 and performing low levels of light therapy 81 can all help intervene in the direction of stem cell differentiation after printing.
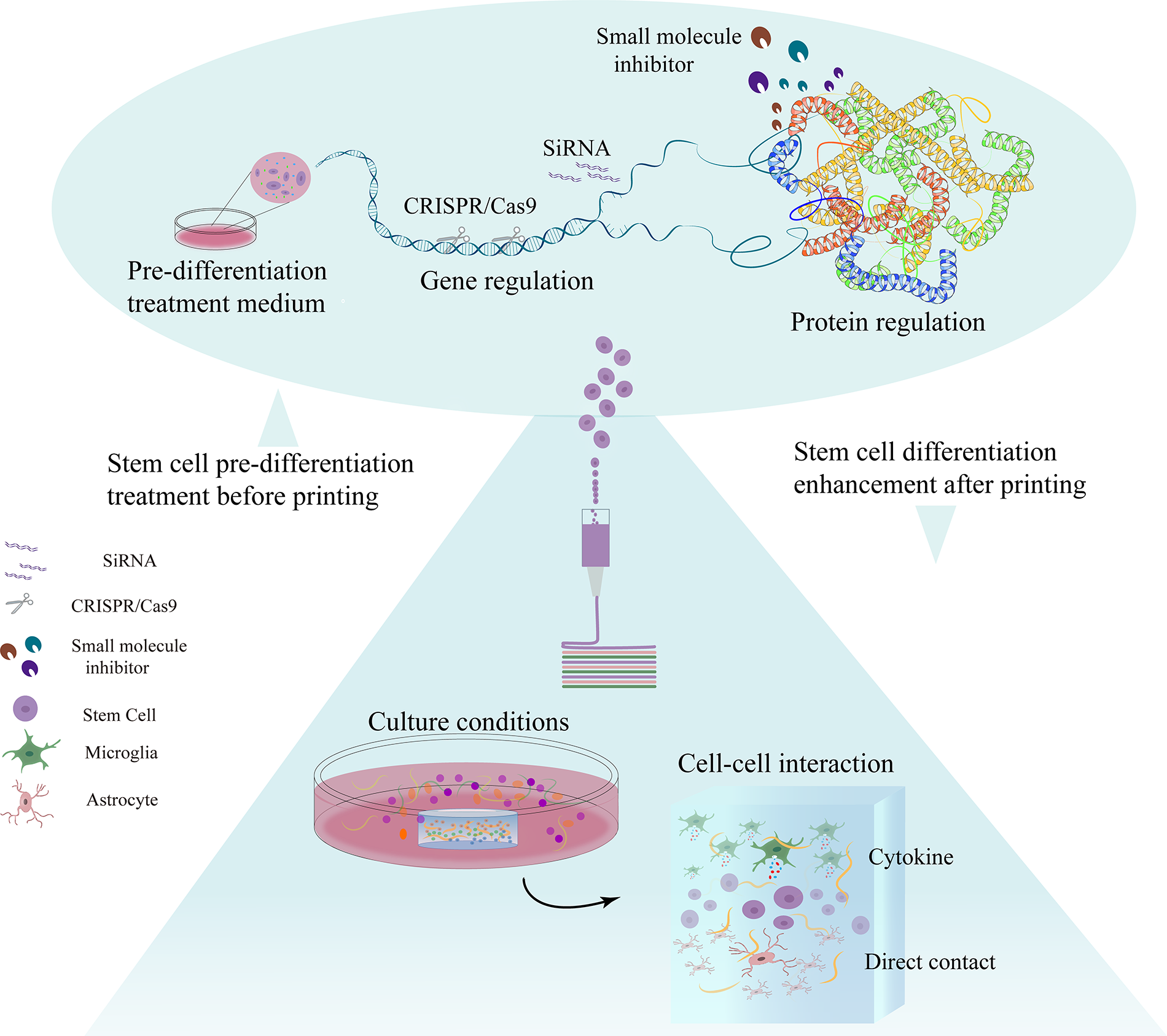
Strategies to induce stems cell differentiation in multicellular neural 3D-bioprinting. Common approaches can be generally categorized into preprinting treatments (top) and postprinting conditions (bottom). Preprinting strategies include gene editing, SiRNA interference, and small molecule inhibitors, aiming to modify protein expression and induce preprinting stem cell differentiation. Postprinting protocols such as culture conditions and cellular compositions can also be used to promote stem cell differentiation after printing.
Disease Models Based on Multicellular Neural Constructs
Compared with 3D bioprinting with a single-cell type, multicellular 3D bioprinting allows for the incorporation of intercellular interactions, thereby providing more physiologically relevant in vitro models for neuroscience research. This encompasses fundamental inquiries such as neuronal migration, development, and neural network formation. Additionally, it includes investigations into related diseases such as neurodegenerative disorders, brain tumors, and cerebrovascular pathologies. These conditions similarly involve complex interplay among diverse cell populations, thus demanding research models that could recapitulate such interactions in vitro.
Neural network construction
The construction of excellent neural network models employing 3D bioprinting should meet the following criteria: the presence of structural hierarchies supporting synapse extension; the inclusion of diverse neuronal subtypes; the maintenance of cellular activity over an extended period of time in vitro; and the demonstration of good electrical conductivity. Currently, the construction of neural networks via 3D bioprinting remains in its nascent stages.23,24 To generate functionally active neural networks within printed constructs, factors such as fundamental printing parameters, 73 axonal outgrowth, synapse formation,101,102 and intercellular interactions, have all to be considered. This section provides a summary of the role of intercellular interactions in neural network assembly in 3D bioprinting.
In the study conducted by Yao et al., the research team employed a GEL-norbornene poly(ethylene glycol)-dithiol (GelNB-PEGdiSH) bioink encompassing rat primary cortical neurons and astrocytes to fabricate cortical structures in vitro. 20 The construct exhibited a calcium transient pattern similar to that of the primary mouse cortex. The authors observed a facilitating effect of glial cells on neurite outgrowth and migration. The neuronal cell types that are predominantly found in the cerebral cortex can be broadly classified into two main categories: GABAergic interneurons and glutamatergic neurons. To more accurately simulate neuronal activities in the cerebral cortex, Yan et al. developed a neural network model utilizing a fibrin gel (2.5 mg/mL fibrinogen + 0.5 U thrombin) mixed with collagen hydrogel, which supports the growth of diverse neuronal cell subtypes and glial cells. 25 Initially, the team injected a 1:4 mixture of medial ganglion eminence-derived GABA and cortical (glutamatergic) progenitor cells into the structure to ascertain whether the cells could differentiate correctly. Over the subsequent 60 days, the team observed that the two types of progenitor cells underwent the requisite differentiation into distinct neuronal subtypes and exhibited stable expression of relevant markers. Additionally, an increase in spontaneous action potentials of the neuronal cells, the formation of synaptic structures, and the formation of synapses were observed in this printed structure approximately 20 days after printing, which occurred earlier than in 2D cultures.25,103 Subsequently, the team introduced astrocytes into the printed structure at a 1:1 ratio. The astrocytes were integrated into the printed structure and correctly recycled neurotransmitters released by neurons in the tissue. 104
Jin et al. employed 3D printing with preprocessed iPSCs for the construction of a cortex-like model in vitro. 105 The team employed 3D droplet printing based on the ECM to construct a six-layer cortex-like printed structure. The structure comprised two principal cell types: upper layer neuronal progenitor cells and deep layer neuronal progenitor cells (DNPs), both of which were derived from iPSCs. During the subsequent 8 weeks, the structure gradually showed a layered distribution similar to that of cortical organoids. 26 After implanting the printed structures into the mouse cortex, it was observed that printed DNPs generated similar calcium oscillation frequencies with the host neurons, indicating a possibility of synaptic connections between them.
In the study conducted by Koch et al., NSCs were more electrophysiologically active when they were printed with astrocytes. 94 This enhancement was found to vary with the percentage of astrocytes present, with an increasing trend observed in the 20–50% range. However, this effect was not observed in 3D coculture of astrocytes and neurons, which may be related to the fact that NSCs can differentiate into other glial cells.
Beyond glial cells, some recent studies have identified a potentially positive role for ECs in neuronal network formation. Abdelrahman et al. utilized 3D bioprinting to create a 3D coculture of HUVECs, dopaminergic (DA) neurons, and cortical neurons. They observed a denser neurite network compared with individual printing. 106
Disease modeling and applications
3D printing is being used in the construction of disease models, including epilepsy, 107 brain injury, 108 the blood–brain barrier defect,109,110 and myelin sheath degeneration. 111 To accurately replicate the complex nature of disease occurrence and progression, it is vital to incorporate a diverse range of cell types in a 3D ex vivo setting (Table 3).
3D Printed Models of Neurological Diseases

AD, Alzheimer’ disease; GO, graphene oxide; GSCs, glioma stem cells; IVFK, Ac-Ile-Val-Phe-Lys-NH2; IVZK, Ac-Ile-Val-Cha-Lys-NH2; PD, Parkinson’s disease; SA, sodium alginate; SISMA, methacrylate-modified decellularized small intestinal submucosa; VM DA, ventral midbrain dopaminergic human.
Abbreviated Table

Neurodegenerative diseases
Parkinson’s disease (PD) and Alzheimer’s disease (AD) are hallmark examples of neurodegenerative disorders. These diseases are characterized by irreversible damage to the structural and functional integrity of the human brain, ultimately leading to compromised behavioral and cognitive functions. Despite extensive research, the precise pathologies of these diseases largely remain elusive. Utilizing in vitro disease models for investigation stands as one of the most cost-effective approaches. However, due to the inherent complexity of the brain and current technological limitations, the construction of accurate in vitro disease models remains challenging. To this end, 3D bioprinting offers a distinctive route for establishing disease models. 120 Through multicellular 3D bioprinting, researchers gain increased flexibility in observing the roles played by distinct cell types within these disease-relevant constructs.
PD progression is marked by the loss of DA neurons in the substantia nigra, prompting many in vitro model designs to center on recapitulating DA neuron depletion. In multicellular bioprinting, the foremost consideration remains compatibility between printing parameters for different cell types. In this regard, Abdelrahman et al. chose self-assembling short peptides as hydrogel material, which proved compatible with both DA neuron and ECs in terms of survival and growth.
106
Both in monocultures and 3D coculture, DA neurons grown on this peptide substrate displayed superior survival rates and ATP release compared with those cultured on poly-
Besides DA neuron loss, many other factors are equally crucial in the pathogenesis of PD, including the involvement of the immune system. Rueda-Gensini et al.’s PD model specifically focuses on the immune-pathological aspects. This model integrates astrocytes and microglia into a 3D coculture system and then exposes the system to α-synuclein to elicit PD-like neuroimmune pathology. 113 Notably, this model did not simply mix the three cell types. Instead, the entire 3D coculture was divided into two compartments. In one compartment, astrocytes and microglia were directly 3D cocultured, while in the other compartment, DA neurons were spatially segregated. With such arrangement cells interacted solely through paracrine signaling, thereby simulating the microenvironmental impact on PD. 113 The model’s validity was substantiated through assessments of DA neuron apoptosis, mitochondrial dysfunction, α-synuclein distribution, and changes in key DA-related genes (TH expression downregulated approximately 64-fold, KCNJ6 and DAT downregulated about 2000-fold, Syn1 downregulated approximately 1000-fold, and PINK1 and Parkin expression decreased). 113 Compared with alternative 3D models like organoids, this coculture system manifests PD characteristics at an earlier stage (3D bioprinting system: 7 days; organoids: 21 days).113,122
AD, a progressive neurodegenerative disorder, exhibits a strong positive correlation between its incidence and advancing age, with its most characteristic features being the accumulation of β-amyloid plaques and neurofibrillary tangles resulting from excessive phosphorylation of tau protein. 123 Consequently, when establishing in vitro models, these pathognomonic hallmarks often serve as criteria for assessing model validity. 124 AD models can be established through various approaches, such as overexpression or transfection of Aβ-related protein genes, 125 direct incorporation of relevant peptides like Aβ1-42, 126 or utilization of patient-derived iPSCs that differentiated into neurons.116,127 Researchers have exploited these 3D models to investigate AD from metabolic, 125 pathophysiological,116,126 and other perspectives. Current AD models constructed using 3D bioprinting typically use a single type of cell, for example, hiPSC 116 or SH-SY5Y. 128 With ongoing technological advancements, multicellular bioprinting holds promise to augment existing AD models by incorporating microglia or astrocytes given their intimate association with AD-related neuroinflammation. 123
Neoplasms
Glioblastoma (GBM) is the most prevalent primary malignant brain tumor in adults, with an overall incidence rate of 3.2 per 100,000 individuals. 129 Characterized not only by its high prevalence, GBM is also marked by unclear pathological factors and rapid recurrence, typically manifesting within approximately 8 months postsurgery. 130 Compared with the 2D model, the 3D model is shown to yield better cell survival and electrophysiological activity, making it more suitable for disease research. 101 We herein summarize the current progress in the development of in vitro 3D GBM models utilizing multicellular bioprinting.
The tumor microenvironment (TME) of GBM in humans typically involves a complex interplay of tumor-associated macrophages, glial cells, neurons, MSCs, and perivascular cells. 131 In studying GBM, Bai et al. have modeled the in vitro interaction between GBM and neurons. Their team utilized 3D bioprinting technique to design a construct consisting of an outer hemisphere shell containing neurons and an inner hemisphere harboring glioma cell, achieved by separately printing the two cell types and subsequently assembling them. Notably, compared with monoculture models, the GBM coculture model demonstrated significantly higher cell viability (GL261/neuron: 70.24% ± 2.78%, neuron: 60.00% ± 2.76%). 117 Furthermore, Bai et al. observed, via calcium imaging, that neurons in close proximity to glioma cells exhibited heightened Ca2+ fluorescence intensity, indicative of increased electrical signaling activity in these neurons. This finding began to reveal the intricate interplay between neurons and glioblastoma cells, suggesting that in the early stages of the disease, glioma cells may promote axonal and synaptic growth in neurons, leading to enhanced neuronal electrical activity. 132 Correspondingly, this facilitatory effect also favors GBM progression. As neuronal activity intensifies, elevated Ca2+ concentrations can activate ERK/MAPK and PKA signaling pathways, promoting glioma cell migration and invasion, 117 thereby exacerbating disease progression.
In tumors, ECs play a vital role in shaping the vasculature of the TME, closely intertwined with the progression of GBM. 133 The model developed by Marino et al. revolves around 3D coculturing ECs with GBM cells, employing two-photon lithography technology to create an intriguing and versatile 3D coculture system that allows different cell types to be assembled like building blocks. Specifically, they encapsulate glioblastoma cells within larger dodecahedron-shaped scaffolds that serve as the core. Then they incorporate healthier cells, such as ECs and neurons, into smaller tetrahedral scaffolds to function as modular components. These scaffolds then self-assemble magnetically due to the magnetic properties in their design, ultimately achieving the goal of cocultivation. 48 Using this 3D coculture model, Marino et al. confirm the facilitatory effect of ECs on GBM migration, consistent with previous findings. 134 Additionally, Cho et al. used glioblastoma with ECs for multicellular bioprinting 135 and introduced the hypoxic environment prevalent in solid tumors. 136 Comparative analysis with conventional modeling regions reveals that areas featuring gradient oxygen exhibit accelerated expression of cancer-related genes, thereby abbreviating the modeling time. Using this model, the team recapitulates certain clinical responses observed in patients following antitumor drug treatment, such as drug resistance, thereby providing a preliminary validation for the model’s reliability. 135
In pursuit of a more faithful representation of the TME, Neufeld et al. incorporated microglia and astrocytes, alongside ECs and pericytes, into a multicomponent bioprinting framework. 137 The GBM disease model constructed from these five cell types displays gene alterations similar to those seen in murine models under disease conditions, changes not observed in 2D models. Genes exhibiting such alterations include MMP9 (facilitating tumor angiogenesis), IRF-1, and SEMA7A (involved in tumor migration and invasion). Compared with 2D models, 3D GBM models demonstrate heightened drug resistance, potentially better reflecting the in vivo drug tolerance of tumors, and making them more suitable as platforms for drug screening.137,138
Current research has firmly established the profound involvement of macrophages in GBM progression and recurrence.139–141 Herinrich et al. have contributed to this line of inquiry using 3D bioprinting technology, establishing an immune-inclusive GBM model through 3D coculture of macrophages with glioblastoma cells. 142 Their team first observed that GBM cells within the 3D construct exhibit a heightened ability to recruit macrophages compared with 2D models and single-component systems. Additionally, utilizing this model, Herinrich et al. evaluated the efficacy of carmustine, Stat6 inhibitors, and colony-stimulating factor 1 receptor (Csf-1r) inhibitors on macrophages and GBM cells. The 3D coculture model demonstrated a significantly higher overall drug tolerance (2D [IC50]: 139 µm, 3D [IC50]: 581 µm). 142 This finding aligns with results from Hermida et al., who constructed a GBM model by coprinting glioblastoma cells, macrophages, and FIB. 143 In their model, GBM cells and G7 cells displayed approximately twofold increases in their temozolomide IC50 values when compared with 2D culture (2D: 928.7 ± 1.0 µM and 333.3 ± 1.1 µM; 3D: 1994 ± 1.0 µM and 748.8 ± 1.1 µM). The addition of macrophages exacerbated this trend. Macrophages incorporated into the 3D coculture system decreased GBM cell sensitivity to the drug and raised their survival rate from 50.1% ± 6.7% to 73.8% ± 5.2%. 143 Collectively, these outcomes substantiate the notion that the interactions between macrophages and GBM cells foster tumor progression. 140
Significant progress has been made in the construction of multicellular disease models for GBM. 3D bioprinted ex vivo disease models hold considerable potential for advancement. Key areas for improvement include expanding the repertoire of cell types incorporated in the printing process and prolonging the viability of cells within the models, both of which are crucial for conducting long-term observations.
Discussion and Prospects
In neuroscience, it has been widely suggested that single-cell types frequently fail to adequately represent complex physiological or pathological states. Consequently, bioprinting has evolved from using only one cell type to encompass multiple cell types. Significant advances have been made in the field of multicellular neural tissue bioprinting.135–138 Of particular note is the exploration of interactions between neural cells and cells from nonneural tissues, including brain vessels and the blood–brain barrier, as well as interactions between neural cells and muscle cells.16,17,18
In relation to printing materials, further exploration is required for composite inks or scaffolds tailored to different types of neural cells. Given the low stiffness of neural tissue, it is crucial to consider how to balance the integrity of the printed structure with postprinting cell survival, attachment, and extension.
In relation to current neural tissue models, the printing of high-level structures such as the cortex and blood vessels remains in its nascent stages, exhibiting limited functionality. The vascular network is a pivotal component of the blood–brain barrier, and constructing a functional blood–brain barrier would facilitate drug screening processes. It is imperative to ensure that the proportion of different cell types within the printed structures mirrors the in vivo situation for accurate drug screening.
In addition to the multicellular types of cell printing discussed in this review, 3D bioprinting offers considerable room for further exploration. From an application perspective, this includes optimizing 3D bioprinting for clinical use,51,56 refining models for high-throughput drug screening, and incorporating external stimuli into printed structures. 118 From a printing protocol perspective, the introduction of in situ printing or a multijet printing system may help to better print multicellular neuronal cells.
Advantages and disadvantages of 3D bioprinting in comparison to other 3D culture techniques, as well as the mutual enhancements between them have been discussed.2,4,121 For instance, the combination of microfluidics with 3D bioprinting can address the limitations of 3D bioprinting in establishing the blood–brain barrier.30–32 Alternatively, the development of particulate hydrogels can be achieved through the use of microfluidics, which offers precise control over particle size and shape. 33 In the context of organoids, the application of 3D bioprinted blood vessels in organoid models can mitigate central ischemic necrosis, which is frequently observed in the long-term culture of organoids.16,34 The employment of extrusion-based 3D bioprinting technology to construct organoid models has been shown to effectively address the issues of poor reproducibility and low throughput associated with conventional organoid fabrication methods. 35 The embedded printing method facilitates the direct integration of organoids into 3D bioprinted structures, thereby enhancing the physiologically relevance of the resulting 3D bioprinting models. 35
Declaration of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this work, the author used Qwen for touch-ups. After using this tool, the authors reviewed and edited the content as needed and took full responsibility for the content of the publication.
Authors’ Contributions
Z.L.: Conceptualization, methodology, software, investigation, formal analysis, writing—original draft, and writing—review and editing. T.S.: Data curation and writing—original draft. Y.Y.: Visualization and investigation. H.Z.: Conceptualization, funding acquisition, resources, supervision, and writing—review and editing.
Footnotes
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Funding Information
This work was funded by the Anhui Provincial Natural Science Foundation (2308085MH282 granted to H.Z.).
Data Availability
The data that support the conclusions of this article are all included in this article.